Odstavec předpisu 331/2004 Sb.
Vyhláška Ministerstva zemědělství č. 331/2004 Sb., o opatřeních k zabezpečení ochrany proti zavlékání a šíření původce bakteriální kroužkovitosti bramboru a původce bakteriální hnědé hniloby
Příl.1
Příloha 1
A. METODY DIAGNÓZY, DETEKCE A IDENTIFIKACE PŮVODCE KROUŽKOVITOSTI
Předložené postupové diagramy popisují různé postupy, které jsou součástí:
i) diagnózy kroužkovitosti v hlízách bramboru a rostlinách bramboru,
ii) detekce
Clavibacter michiganensis
subsp. sepedonicus ve vzorcích hlíz
bramboru a rostlin bramboru, iii) identifikace
Clavibacter michiganensis
subsp. sepedonicus
.OBECNÉ ZÁSADY
Optimalizované protokoly pro různé metody, validovaná činidla a podrobnosti
pro přípravu testovaných a kontrolních materiálů jsou uvedeny v dodatcích. Seznam
laboratoří, které se podílely na optimalizaci a validaci protokolů, je v dodatku
č. 1.
Protože protokoly obsahují zjištění karanténního organismu a zahrnují použití
životaschopných kultur
Clavibacter michiganensis
subsp. sepedonicus
jako kontrolních
materiálů, je nutné pracovat za vhodných karanténních podmínek s odpovídajícím zařízením
na odstraňování odpadů a za podmínek příslušných povolení vydaných Ústavem. Testovací
parametry musí zajistit stálé a reprodukovatelné zjištění úrovní Clavibacter michiganensis
subsp. sepedonicus jako stanovené prahy vybraných metod.Zcela nezbytná je příprava pozitivních kontrol. Testování podle požadovaných
prahů také zahrnuje správné nastavení, údržbu a kalibraci zařízení, pečlivé zacházení
s činidly a jejich uchovávání a všechna opatření pro zamezení kontaminace mezi vzorky,
např. oddělení pozitivních kontrol od testovaných vzorků. Musí být uplatněno standardní
řízení kvality, aby se zabránilo administrativním a jiným chybám, zvláště při označování
a v dokumentaci.
Podezření z výskytu, jak je uvedeno v § 4 odst. 1, naznačuje pozitivní výsledek
z diagnostických nebo screeningových testů provedených na vzorku, jak je znázorněno
v níže uvedených postupových diagramech.
Pokud je první screeningový test (IF nebo PCR/FISH) pozitivní, existuje podezření
na infekci původcem kroužkovitosti a musí být proveden druhý screeningový test. Pokud
je i druhý screeningový test pozitivní, pak je podezření z výskytu potvrzeno a musí
se pokračovat v testování podle daného schématu. Pokud je druhý screeningový test
negativní, pak vzorek není považován za infikovaný původcem kroužkovitosti. Z toho
důvodu je pozitivní IF test podle § 4 odst. 1 definován jako pozitivní výsledek IF
testu potvrzený druhým screeningovým testem (PCR/FISH).
Potvrzená přítomnost vyžaduje izolaci a identifikaci čisté kultury
Clavibacter
michiganensis
subsp. sepedonicus
s potvrzením patogenity.1. Použití postupových diagramů
1.1. Postupový diagram pro diagnózu bakteriální kroužkovitosti v hlízách
bramboru a v rostlinách bramboru vykazujících příznaky bakteriální kroužkovitosti
Postup testování je určen pro hlízy a rostliny bramboru s podezřením z výskytu
nebo typickými příznaky kroužkovitosti. Zahrnuje rychlý screeningový test, izolaci
patogenu z infikovaného vodivého pletiva na diagnostickém médiu a v případě pozitivního
výsledku identifikaci kultury
Clavibacter michiganensis
subsp. sepedonicus
.

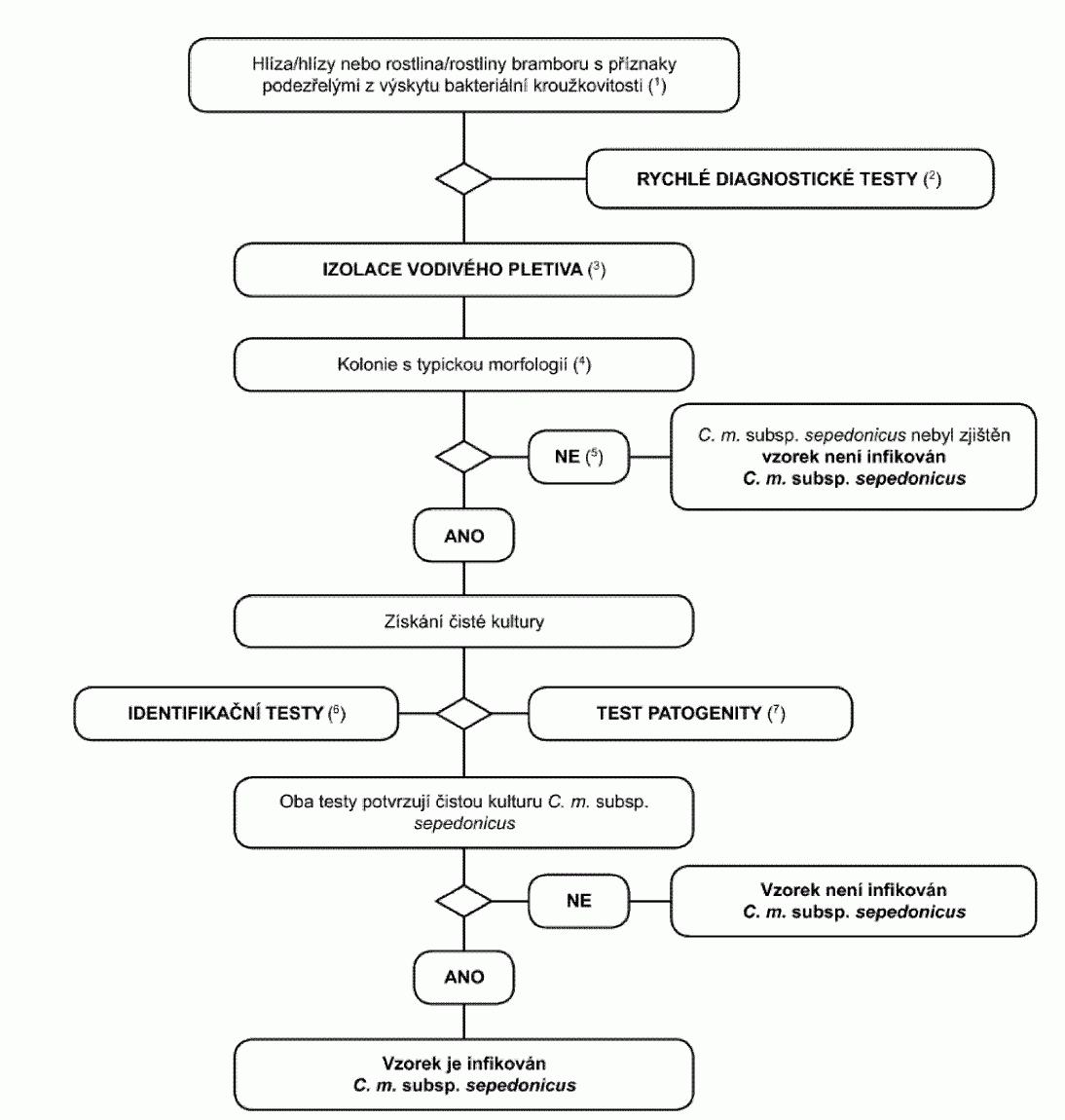
(1) Popis příznaků je v oddíle 2.
(2) Vhodné testy jsou: - IF test (oddíl 4),
- PCR test (oddíl 6),
- FISH
test (oddíl 5).
(3) Ačkoli izolace patogena z rostlinného materiálu s typickými příznaky
roztíráním suspenzí na média není komplikovaná, kultivace v pokročilých stádiích
infekce se nemusí podařit. Saprofytické bakterie, které rostou na infikovaném pletivu,
mohou přerůst nebo potlačovat patogena na izolačním médiu. Proto se doporučuje používat
neselektivní i selektivní média, nejlépe MTNA (oddíl 8) nebo biotest (oddíl 7).
(4) Popis typické morfologie kolonie je v oddíle 8.
(5) Pokud je izolační zkouška negativní, ale příznaky choroby jsou typické,
musí být izolace provedena znovu.
(6) Spolehlivé identifikace čisté kultury Clavibacter michiganensis subsp.
sepedonicus se dosáhne za použití testů uvedených v oddílu 9.
(7) Zkouška patogenity je popsána v oddíle 10.
1.2. Postupový diagram pro diagnózu bakteriální kroužkovitosti ve vzorcích
bezpříznakových hlíz bramboru Postup testování je určen ke zjištění latentních infekcí
v hlízách bramboru. Pozitivní výsledek z nejméně dvou screeningových testů, z nichž
každý je založen na jiném biologickém principu, musí být doplněn izolací patogena,
a následně, v případě izolace typických kolonií, potvrzením, že čistá kultura je
Clavibacter michiganensis
subsp. sepedonicus
. Pozitivní výsledek pouze jednoho screeningového
testu není dostačující k tomu, aby byl vzorek považován za podezřelý. Screeningové a izolační testy musí umožnit detekční práh 10
3
až 104
buněk/ml
resuspendované pelety zahrnutý jako pozitivní kontroly v každé sérii testů.

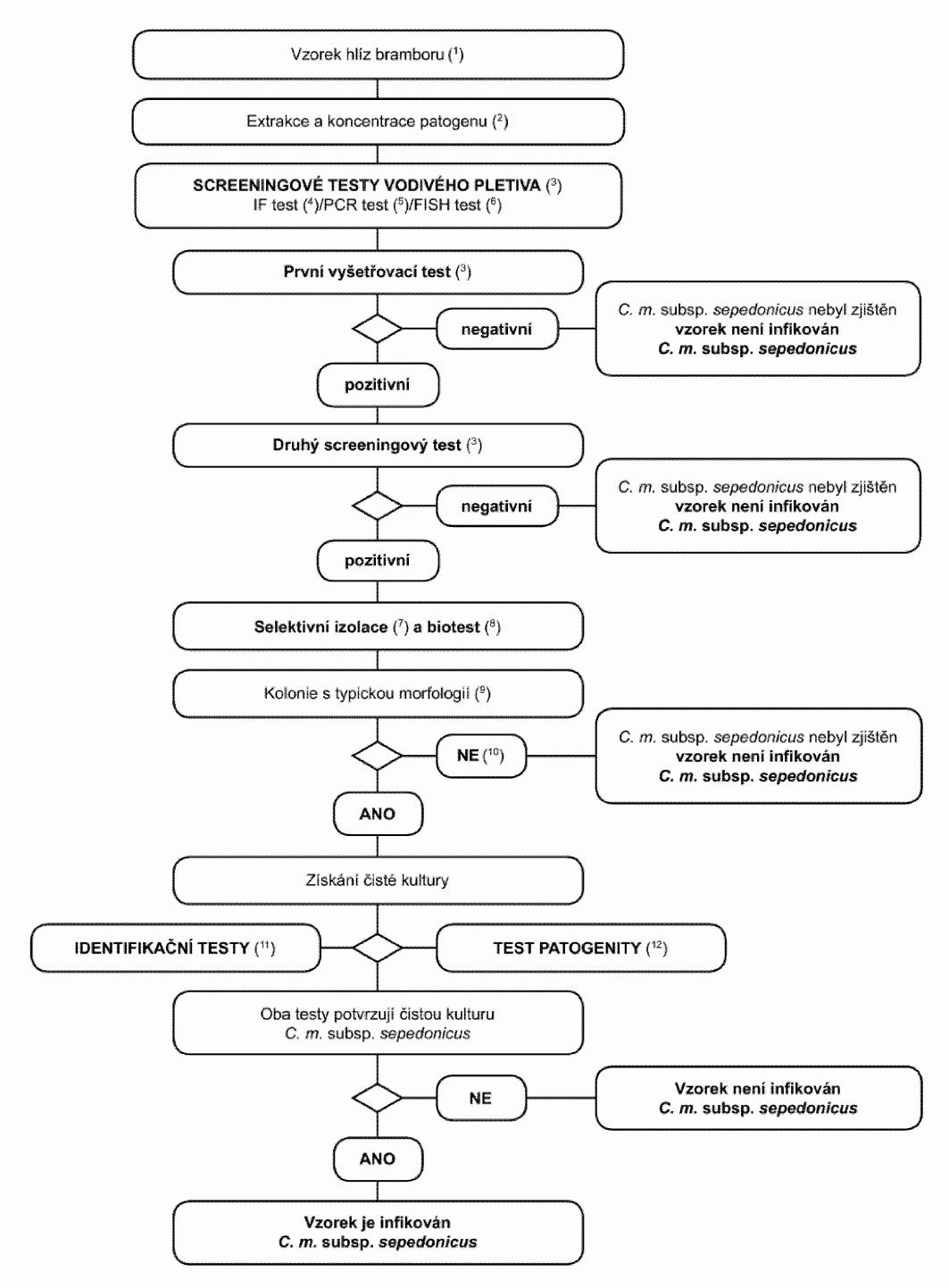
(1) Standardní velikost vzorku je 200 hlíz, ačkoli postup lze použít i na
menší počet, jestliže 200 hlíz není k dispozici.
(2) Metody extrakce a koncentrace patogena jsou popsány v oddílu 3.1.
(3) Pokud jsou alespoň dva testy založené na různých biologických principech
pozitivní, musí být provedena izolace a potvrzení. Provede se nejméně jeden screeningový
test. Pokud je tento test negativní, je vzorek považován za negativní. V případě,
že je tento test pozitivní, je pro ověření prvního pozitivního výsledku nezbytný
ještě jeden nebo více screeningových testů založených na různých biologických principech.
Pokud je druhý nebo další test negativní, je vzorek považován na negativní. Další
testy nejsou nutné.
(4) Test imunofluorescence (IF).
Pro vyšetření IF se vždy použije polyklonální
protilátka, další monoklonální protilátky umožní větší přesnost (viz oddíl 4).
(5) PCR test.
Použijí se vhodná validovaná PCR činidla a protokoly (viz
oddíl 6).
(6) FISH test.
Použijí se validovaná činidla a protokoly (viz oddíl 5).
(7) Selektivní izolace.
Toto může být v mnoha případech vhodná metoda pro
přímou izolaci Clavibacter michiganensis subsp. sepedonicus za použití MTNA média
nebo NCP-88 média a ředění resuspendované pelety 1/100. Typické kolonie lze získat
během 3-10 dní po rozetření na médium. Kulturu patogena lze následně vyčistit a identifikovat.
Pro úplné využití potenciálu test vyžaduje opatrnou přípravu pletiva z pupkové části,
aby se omezily sekundární bakterie, které jsou konkurencí Clavibacter michiganensis
subsp. sepedonicus na médiu a které mohou patogena přerůst. Pokud se kultivační metoda
takto nepodaří, musí být izolace provedena z rostlin použitých pro biotest (viz oddíl
8).
(8) Biotest se používá k izolaci Clavibacter michiganensis subsp. sepedonicus
z pelet bramborových extraktů pomocí selektivního obohacení v rostlinách lilku vejcoplodého
(Solanum melongena). Test vyžaduje optimální inkubační podmínky stanovené pro tuto
metodu. Bakteriální inhibitory Clavibacter michiganensis subsp. sepedonicus na MTNA
nebo NCP-88 médiu pravděpodobně tento test narušovat nebudou (viz oddíl 7).
(9) Typická morfologie kolonie je popsána v oddílu 8.
(10) Kultivace nebo biotesty mohou selhat z důvodů konkurence nebo inhibice
saprofytickými bakteriemi. Pokud jsou výsledky screeningových testů pozitivní, ale
izolační testy negativní, opakují se izolační testy ze stejné pelety nebo dodatečným
odebráním vodivého pletiva z blízkosti pupkového konce hlíz stejného vzorku a v případě
potřeby se provede test dalších vzorků.
(11) Spolehlivá identifikace čistých kultur podezřelých na Clavibacter michiganensis
subsp. sepedonicus se dosáhne použitím testů popsaných v oddílu 9.
(12) Test patogenity je popsán v oddílu 10.
1.3. Postupový diagram pro diagnózu bakteriální kroužkovitosti ve vzorcích
bezpříznakových rostlin bramboru

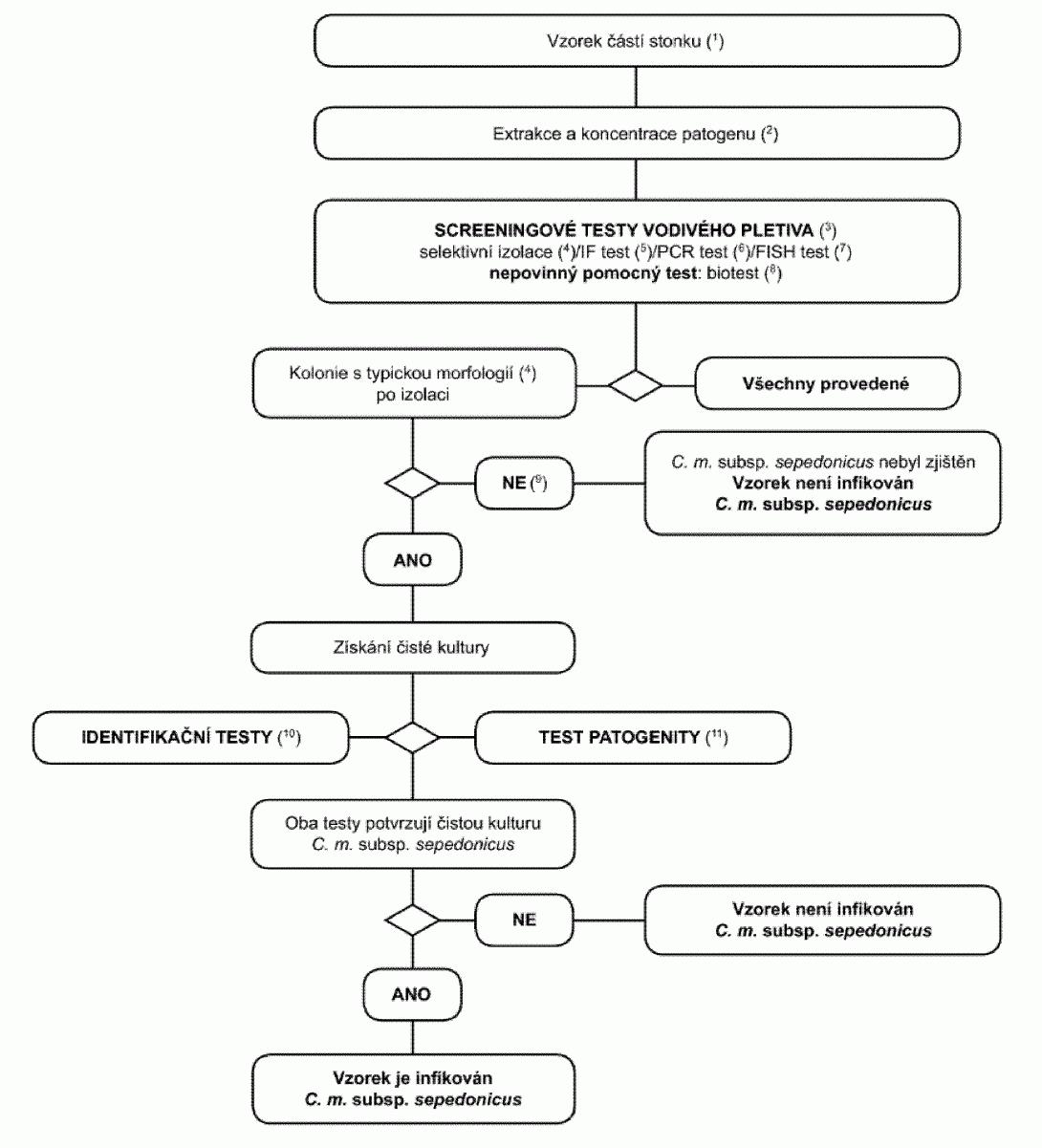
(1) Doporučené velikosti vzorků - viz oddíl 3.2.
(2) Metody extrakce a koncentrace patogena jsou popsány v oddílu 3.2.
(3) Pokud jsou alespoň dva testy založené na různých biologických principech
pozitivní, musí být provedena izolace a potvrzení. Provede se nejméně jeden screeningový
test. Pokud je tento test negativní, je vzorek považován za negativní. V případě,
že je tento test pozitivní, je pro ověření prvního pozitivního výsledku nezbytné
provedení druhého nebo více screeningových testů založených na různých biologických
principech. Pokud je druhý nebo další test negativní, je vzorek považován za negativní.
Další testy nejsou nutné.
(4) Selektivní izolační test a typická morfologie kolonie jsou popsány v
oddílu 8.
(5) IF test je popsán v oddílu 4.
(6) PCR test je popsán v oddílu 6.
(7) FISH test je popsán v oddílu 5.
(8) Biotest je popsán v oddílu 7.
(9) Kultivace nebo biotest mohou selhat z důvodů konkurence nebo inhibice
saprofytickými bakteriemi. Pokud jsou výsledky screeningových testů pozitivní, ale
izolační testy jsou negativní, opakují se izolační testy a v případě potřeby se provede
test dalších vzorků.
(10) Spolehlivé identifikace čistých kultur, u kterých je podezření, že se
jedná o Clavibacter michiganensis subsp. sepedonicus, se dosáhne pomocí testů popsaných
v oddílu 9.
(11) Test patogenity je popsán v oddílu 10.
2. Vizuální vyšetření na přítomnost příznaků kroužkovitost
i2.1. Rostliny bramboru
V evropských klimatických podmínkách se příznaky na poli zjistí jen zřídkakdy
a často pak až na konci sezóny. Kromě toho jsou příznaky skryté nebo se zamění s
příznaky jiných chorob, stáří nebo mechanických poškození. Proto mohou být příznaky
při kontrole na poli snadno přehlédnuty. Příznaky vadnutí jsou velmi odlišné od příznaků
hnědé hniloby; vadnutí je obvykle pomalé a zpočátku omezené na okraje listů. Mladé
infikované listy často i přes infekci nadále rostou, i když růst v infikovaných místech
je omezený. Tím vznikají neobvykle tvarované listy. Listy postižené ucpáním vodivých
pletiv na spodní části stonku často mají chlorotické, žluté až oranžové mezižeberní
partie. Infikované děložní lístky, listy a dokonce stonky mohou eventuálně odumřít.
Často jsou listy a hlízy pouze menší. Příležitostně jsou rostliny zakrnělé. Barevné
snímky řady příznaků jsou na internetové stránce http://forum.europa.eu.int/Public/irc/sanco/Home/main.
2.2. Hlízy bramboru
Nejranějšími příznaky jsou slabá sklovitost nebo průsvitnost pletiva bez
měknutí v okolí cévního systému, zejména blízko pupku. Prstenec svazků cévních na
pupkovém konci hlízy může mít nepatrně tmavší zbarvení než obvykle. Prvním dobře
identifikovaným příznakem je žlutavé zabarvení prstence cévních svazků a stav, kdy
při jemném zmáčknutí hlízy vystupují z cév sloupky sýrovité hmoty. Tento exsudát
obsahuje miliony bakterií. Vodivá pletiva mohou zhnědnout a příznaky na hlízách jsou
v tomto stádiu podobné příznakům hnědé hniloby způsobené Ralstonia solanacearum.
Zpočátku mohou být tyto příznaky omezeny na jednu část prstence, nemusí se vyskytovat
jen blízko pupkové části a mohou se postupně šířit na celý prstenec. S postupem infekce
dochází k destrukci vodivých pletiv: vnější korová část se může oddělit od vnitřní
korové části. V pokročilých stadiích infekce se objevují na povrchu hlízy praskliny,
často s červenohnědými okraji. V poslední době se v Evropě objevilo několik případů,
kdy střední kůra hnije s prstencem svazků cévních, čímž dochází k druhotnému poškození
se vznikem vnitřních dutin a nekrózy. Druhotná houbová nebo bakteriální infekce může
příznaky maskovat a může být obtížné, ne-li nemožné, rozlišit pokročilé příznaky
kroužkovitosti od jiných hnilob bramboru. Možné jsou i atypické příznaky. Barevné
snímky řady příznaků jsou na internetové stránce http://forum.europa.eu.int/Public/irc/sanco/Home/main.
3. Příprava vzorků
3.1. Hlízy bramboru
Poznámka:
- Standardní velikost vzorku je 200 hlíz na 1 test. Intenzivnější
vzorkování vyžaduje více testů na vzorcích této velikosti. Větší množství hlíz ve
vzorku vede k zpomalení nebo složitějšímu výkladu výsledků. Postup však lze vhodně
použít i pro vzorky s méně než 200 hlízami, pokud je k dispozici menší množství hlíz.
-
Validace všech níže uvedených zjišťovacích metod je založená na testování vzorků
o velikosti 200 hlíz.
- Níže popsaný bramborový extrakt lze použít také pro zjištění
původce hnědé hniloby,
Ralstonia solanacearum
. Nepovinné ošetření před přípravou vzorku:
Hlízy se omyjí. Použijí se vhodné
dezinfekční prostředky (s obsahem chlóru, jestliže má být proveden PCR test, pro
odstranění možné patogenní DNA) a mycí prostředky mezi každým vzorkem. Hlízy se nechají
oschnout na vzduchu. Tento postup mytí je obzvlášť užitečný v případě, že je ve vzorku
příliš zeminy a jestliže se má provádět PCR test nebo přímá izolace.
3.1.1. Pomocí čistého a dezinfikovaného skalpelu nebo nožem či škrabkou
na brambory se odstraní slupka na pupkovém konci každé hlízy. Opatrně se vyříznou
konické výkrojky vodivého pletiva z pupkových konců brambor. Na minimum se omezí
nadbytečná část pletiva nezahrnující cévní svazky. Po odebrání musí být výkrojky
z pupkových konců zpracovány do 24 hodin nebo konzervovány při - 20°C po dobu nejdéle
dvou týdnů.
Všechny hlízy s podezřelými příznaky bakteriální kroužkovitosti se dají
stranou a testují se odděleně. Pokud jsou při vyříznutí výkrojku z pupkového konce
zjištěny příznaky kroužkovitosti, provede se vizuální vyšetření této hlízy po naříznutí
hlízy na pupkovém konci. Všechny naříznuté hlízy s podezřelými příznaky se nechají
korkovatět po dobu 2 dnů při pokojové teplotě a uchovávají se v karanténě při teplotě
4 - 10 °C až do ukončení všech testů. Všechny hlízy ve vzorku se uchovávají podle
přílohy č. 2.
3.1.2. Výkrojky z pupkové části se shromáždí v nepoužitých nádobách na jedno
použití, které jsou uzavíratelné a/nebo utěsnitelné (v případě, že jsou nádoby znovu
používány, musí být důkladně vyčištěny a dezinfikovány prostředky s obsahem chlóru).
Nejlépe je zpracovat výkrojky z pupkového konce okamžitě, pokud to není možné, uchovávají
se v nádobě bez přidání pufru, v chladu po dobu maximálně 72 hodin nebo při pokojové
teplotě maximálně 24 hodin. Schnutí a suberizace výkrojků a růst saprofytů v průběhu
skladování může bránit zjištění přítomnosti bakterie způsobující kroužkovitost.
3.1.3. Výkrojky z pupkového konce se zpracují jedním z následujících postupů:
a) výkrojky se zalijí dostatečným množstvím (přibližně 40 ml) extrakčního
pufru (dodatek 3) a třepají se v rotační třepačce (50-100 ot/min) po dobu 4 hodin
při teplotě nižší než 24 °C nebo po dobu 16-24 hodin chlazené;
nebo
b) výkrojky se homogenizují s dostatečným množstvím (přibližně 40 ml)
extrakčního pufru (dodatek 3), buď v mixéru nebo rozdrcením v utěsněném jednorázovém
maceračním sáčku za použití gumového tloučku nebo vhodného mlecího zařízení.
Poznámka:
Při homogenizaci vzorků za použití mixéru existuje vysoké nebezpečí
křížové kontaminace vzorků. Je nutno učinit bezpečnostní opatření pro zamezení vzniku
aerosolu nebo rozlití během extrakce. Pro každý vzorek musí být použita čerstvě sterilizovaná
ostří (nože) a nádoby. Pokud má být použit PCR test, je nutno zamezit přenosu DNA
na nádoby nebo mlecí zařízení, doporučuje se drcení v sáčcích na jedno použití a
použití zkumavky na jedno použití.
3.1.4. Dekantuje se supernatant. Pokud je nadměrně zakalený, pročistí se
buď pomalým odstředěním (při maximálně 180 g po dobu 10 minut při teplotě 4-10°C)
nebo vakuovou filtrací (40-100µm), filtr se omyje přídavkem (10 ml) extrakčního pufru
(dodatek 3).
3.1.5. Bakteriální frakce se zahustí odstřeďováním při 7000 g po dobu 15
minut (nebo 10 000 g po dobu 10 minut) při teplotě 4-10°C a odstraní se supernatant,
aniž by se rozvířila peleta.
3.1.6. Resuspenduje se peleta v 1,5 ml peletového pufru (dodatek 3). Použije
se 500 µl pro test na Clavibacter michiganensis subsp. sepedonicus, 500 µl pro Ralstonia
solanacearum a 500 µl pro referenční účely. Přidá se sterilní glycerol na konečnou
koncentraci 10-25 % (v/v) k 500 µl referenční poměrné části a ke zbývající části
vzorku, promíchá se vířením a uloží při teplotě - 16 až -24°C (týdny) nebo při teplotě
-68 až -86°C (měsíce). Extrakty se během testování uchovávají při teplotě 4-10°C.
Opakované zmrazení a rozmrazení se nedoporučuje.
Pokud je nutný transport extraktu,
zajistí se přeprava v chladícím boxu do 24 až 48 hodin.
3.1.7. Je nezbytně nutné, aby se se všemi pozitivními kontrolami a vzorky
Clavibacter michiganensis subsp. sepedonicus zacházelo odděleně, aby se předešlo
kontaminaci. To platí i pro sklíčka na IF testy a všechny testy.
3.2. Rostliny bramboru
Poznámka:
Pro zjištění latentních populací Clavibacter michiganensis subsp.
sepedonicus se doporučuje kontrola kombinovaných vzorků. Postup je vhodný pro kombinované
vzorky až 200 částí stonků. (Pokud se provádí podrobné průzkumy, měly by být založeny
na statisticky reprezentativním vzorku rostlinné populace, která je předmětem vyšetřování.)
3.2.1 Pomocí čistého dezinfikovaného nože nebo zahradnických nůžek se odstraní
část o velikosti 1-2 cm ze spodní strany každého stonku přímo nad povrchem země.
Části stonků se krátce dezinfikují etanolem 70 %, okamžitě osuší savým papírem a
shromažďují v uzavřené sterilní nádobě.
3.2.2. Části stonků se zpracují jedním z následujících postupů:
a) části se zalijí dostatečným množstvím (přibližně 40 ml) extrakčního
pufru (dodatek 3) a třepají v rotační třepačce (50-100 ot/min) po dobu 4 hodin při
teplotě nižší než 24 °C nebo po dobu 16-24 hodin chlazené, nebo
b) části se okamžitě rozdrtí v pevném maceračním sáčku s přiměřeným množstvím
extrakčního pufru (dodatek 3) za použití gumového tloučku nebo vhodného mlecího zařízení.
Pokud to není možné, skladují se části stonků v chladu maximálně 72 hodin nebo maximálně
24 hodin při pokojové teplotě.
3.2.3. Po usazení, které má trvat 15 minut, se dekantuje supernatant.
3.2.4 Další purifikace extraktu nebo koncentrace bakteriální frakce obvykle
není nutná, ale lze ji dosáhnout filtrací a/nebo odstředěním podle popisu v oddílu
3.1.3.-3.1.6.
3.2.5 Čistý nebo koncentrovaný vzorkový extrakt se rozdělí na 2 stejné části,
Jedna polovina se udržuje během testování při teplotě 4-10 °C a druhá polovina se
skladuje s 10-25 % (v/v) sterilního glycerolu při teplotě -16 až -24 °C (týdny) nebo
při teplotě -68 až - 86 °C (měsíce), pokud je nutné další testování.
4. IF test
PRINCIP
Doporučuje se použít IF test jako hlavní screeningový test, protože
byla prokázána jeho schopnost dosáhnout požadovaných prahů.
Použije-li se jako hlavní
screeningový test a jeho výsledek je pozitivní, musí být jako druhý screeningový
test proveden test PCR nebo FISH. Jestliže se test IF použije jako druhý screeningový
test a jeho výsledek je pozitivní, je pro dokončení analýzy nutné další testování
podle postupového diagramu.
Poznámka:
Pokud je IF test používán jako hlavní vyšetřovací test, používá
se vždy polyklonální protilátka. V případě pozitivního výsledku IF testu s polyklonální
protilátkou může být další vyšetření vzorku s monoklonální protilátkou přesnější,
ale méně citlivé.
Používají se protilátky proti známému kmenu původce kroužkovitosti - ATCC33113
(NCPPB 2137) nebo NCPPB 2140. Doporučuje se určovat titr pro každou novou řadu proti
látek. Titr je stanoven jako nejvyšší ředění, při kterém dochází k optimální reakci
při testování suspenze obsahující 10
5
až 106
buněk na 1 ml homologického kmene původce
kroužkovitosti a při použití vhodného konjugátu fluorescenčního isothiokyanatanu
(FITC) podle doporučení výrobce. Nezpracované polyklonální nebo monoklonální protilátky
by měly mít IF titr alespoň 1:2000. Během testu by se měly protilátky používat v
pracovním ředění (WD) v blízkosti titru nebo na titru. Používají se validované protilátky
(viz internetovou stránku http://forum.europa.eu.int/Public/irc/sanco/Home/main). Test se provádí na čerstvě připravených vzorkových extraktech. V případě
potřeby může být úspěšně proveden na extraktech skladovaných při - 68 až - 86 °C
v glycerolu. Glycerol lze ze vzorku odstranit přidáním 1 ml peletového pufru (dodatek
4), opětovným odstředěním po dobu 15 minut při 7 000 g a resuspenzí ve stejném množství
peletového pufru. To často není nutné, zejména pokud jsou sklíčka se vzorky fixována
plamenem (viz 4.2).
Připraví se oddělená pozitivní kontrolní sklíčka s homologickým kmenem nebo
s jiným srovnávacím kmenem původce kroužkovitosti, suspendovaným ve výluhu z brambor
podle dodatku 2 a nepovinně v pufru.
Pletivo infikované přirozenou cestou (uchovávané lyofilizací nebo zmrazením
při teplotě -16 až -24 °C) by se mělo podle možnosti použít jako paralelní kontrola
na stejném podložním sklíčku.
Jako negativní kontroly se použijí alikvotní části vzorkových extraktů,
které byly předtím testovány s negativním výsledkem.
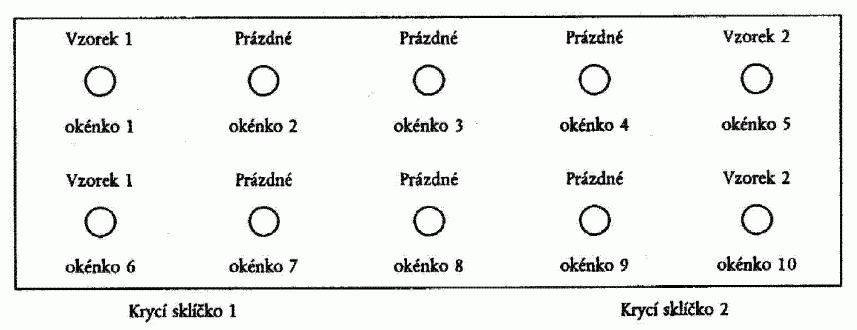
Použijí se mikroskopická sklíčka s více okénky, nejlépe s 10 okénky o průměru
nejméně 6 mm.
Test kontrolního materiálu se provede stejným způsobem jako test vzorků.
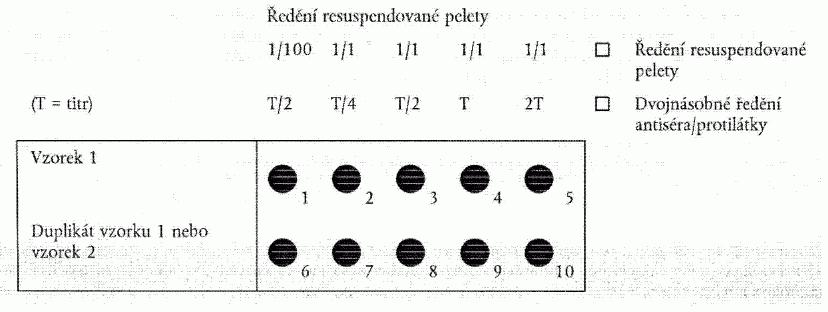
4.1. Sklíčka na test se připraví jedním z následujících postupů:
a) Pro pelety s relativně nízkým množstvím škrobového sedimentu:
Napipetuje
se měřené standardní množství (pro okénka o průměru 6 mm je vhodné 15 µl - pro větší
okénka objem vyšší) roztoku resuspendované bramborové pelety 1/100 do prvního okénka.
Následně se napipetuje stejné množství nezředěné pelety (1/1) do zbývajících okének
v řadě. Druhou řadu lze použít jako duplikát nebo pro druhý vzorek podle obrázku
1.
b) Pro jiné pelety:
Připraví se desetinná ředění (1/10 a 1/100) resuspendované
pelety v peletovém pufru. Napipetuje se měřené standardní množství (pro okénka o
průměru 6 mm je vhodné 15 µl - pro větší okénka objem vyšší) resuspendované pelety
1/100 a každého roztoku do řady okének. Druhou řadu lze použít jako duplikát nebo
pro druhý vzorek podle obrázku 2.
4.2. Vysuší se kapky při pokojové teplotě nebo zahřátím na teplotu 40 až
45 °C. Bakteriální buňky se fixují na sklíčko buď zahřátím (15 minut při teplotě
60 °C), nad plamenem nebo pomocí 95 % etanolu nebo podle zvláštních pokynů dodavatelů
protilátek.
V případě potřeby lze potom fixovaná sklíčka před dalším použitím skladovat
zmrazená v suchém boxu po co možná nejkratší dobu (maximálně 3 měsíce).
4.3. Postup IF testu:
a) Podle přípravy sklíčka na test v oddílu 4.1 písm. a): Připraví se sada
dvojnásobných roztoků protilátky v IF pufru. V první jamce by měl být titr ˝ (T/2),
v ostatních titr Ľ (T/4), titr ˝ (T/2), titr (T) a dvojnásobný titr (2T).
b) Podle přípravy sklíčka na test v oddílu 4.1 písm. b): Připraví se pracovní
ředění (WD) protilátky v IF pufru. Pracovní ředění ovlivňuje přesnost.
Obrázek 1: Příprava sklíčka na test podle oddílu 4.1. písm. a) a oddílu
4.3. písm. a)

Obrázek 2: Příprava sklíčka na test podle oddílu 4.1. písm. b) a oddílu
4.3 b)


4.3.1. Podložní sklíčka se uspořádají na navlhčený papír. Každé testovací
okénko se pokryje kompletně ředěním protilátek. Množství protilátky na každém okénku
musí být nejméně stejné jako množství použitého extraktu.
Pokud nejsou k dispozici konkrétní pokyny od dodavatele protilátek, postupuje
se následovně:
4.3.2. Podložní sklíčka se inkubují na vlhkém papíře přikrytá po dobu 30
minut při pokojové teplotě (18-25 °C).
4.3.3. Setřesou se kapky ze všech podložních sklíček a tato se pečlivě opláchnou
pufrem IF. Umyjí se ponořením po dobu 5 minut v pufru IF Tween (dodatek 3) a následně
v pufru IF. Je třeba zabránit vzniku aerosolu nebo přenosu kapiček, které by mohly
způsobit vzájemnou kontaminaci, a pečlivě odstranit přebytečnou vlhkost jemným osušením.
4.3.4. Sklíčka se umístí na vlhký papír. Testovací okénka se pokryjí ředěním
konjugátu FITC, kterým se stanovuje titr. Množství konjugátu naneseného do okének
musí být stejné jako množství použité protilátky.
4.3.5. Zakrytá sklíčka se inkubují na vlhkém papíru po dobu 30 minut při
pokojové teplotě (18-25 °C).
4.3.6. Kapky konjugátu se setřesou ze sklíček a tato se opláchnou a umyjí
jako předtím (4.3.3.).
Opatrně se odstraní přebytečná vlhkost.
4.3.7. Napipetuje se 5-10 µl 0,1M fosfátového pufru s glycerolem (dodatek
3) nebo komerční krycí tekutiny do každého okénka a přiloží krycí sklíčko.
4.4. Vyhodnocení IF testu
4.4.1. Testovací sklíčka se prohlížejí epifluorescenčním mikroskopem s filtry
vhodnými pro excitaci FITC pod olejovou nebo vodní imersí při zvětšení 500x až 1000x.
Zkoumají se okénka ve dvou navzájem kolmých průměrech a kolem obvodu. U vzorků s
žádnými nebo malým počtem buněk se zkoumá nejméně 40 polí mikroskopu. Nejdřív se
zkontroluje pozitivní kontrolní vzorek. Buňky musí být jasně fluoreskující a zcela
obarvené v určeném titru protilátky nebo pracovním ředění. Pokud je barevnost odchylná,
musí být test IF opakován (oddíl 4).
4.4.2. Pozorují se jasně fluoreskující buňky s charakteristickou morfologií
Clavibacter michiganensis subsp. sepedonicus v testovacích okénkách sklíčka (viz
internetová stránka http://forum.europa.eu.int/Public/irc/sanco/Home/main). Intenzita
fluorescence musí být při porovnání s pozitivním kontrolním kmenem ve stejném ředění
protilátky stejná nebo lepší. Buňky s neúplným zbarvením nebo slabou fluorescencí
nelze brát v úvahu. Při podezření z jakékoli kontaminace musí být test zopakován.
To se může stát, když všechna sklíčka ve skupině vykazují pozitivní buňky díky kontaminaci
pufru nebo při zjištění pozitivních buněk (mimo okénka sklíček) na povrchu sklíček.
4.4.3. Existuje několik problémů podstatných pro přesnost imunofluorescenčního
testu. V peletách z pupkové části bramboru a částí stonku se mohou vyskytnout doprovodné
populace fluoreskujících buněk s atypickou morfologií a křížově reagující saprofytické
bakterie s velikostí a morfologií podobnou Clavibacter michiganensis subsp. sepedonicus.
4.4.4. Berou se v úvahu pouze fluoreskující buňky s typickou velikostí a
morfologií v titru nebo pracovním ředění protilátek podle oddílu 4.3.
4.4.5. Interpretace výsledku IF testu:
a) Při zjištění jasně fluoreskujících buněk s typickou morfologií se odhadne
průměrný počet typických buněk v 1 mikroskopickém poli a vypočítá počet typických
buněk na 1 ml resuspendované pelety (dodatek 4). Výsledek IF je pozitivní u vzorků,
kde je počet typických buněk na 1 ml resuspendované pelety nejméně 5x10
3
. Vzorek
je považován na potenciálně infikovaný a je povinné další testování. b) Výsledek IF testu je negativní pro vzorky, které obsahují méně než
5x10
3
buněk na 1 ml resuspendované pelety a vzorek se považuje za negativní. Další
testování není nutné.5. Test FISH
PRINCIP
Když se jako první screeningový test použije FISH test a je pozitivní,
musí být jako druhý povinný screeningový test proveden IF test. Když se FISH test
provede jako druhý screeningový test a je pozitivní, je k dokončení diagnózy nutné
další testování podle postupového diagramu.
Poznámka:
Používají se validované oligosondy specifické pro Clavibacter
michiganensis subsp. sepedonicus (dodatek 7). Úvodní testování touto metodou by mělo
umožnit reprodukovatelné zjištění alespoň 10
3
-104
buněk Clavibacter michiganensis
subsp. sepedonicus na ml přidané do extraktů ze vzorku, které byly předtím testovány
s negativním výsledkem. Následující postup by měl být pokud možno proveden s čerstvě
připravenými extrakty, ale je možné jej úspěšně provést s extraktem, který byl uchován
v glycerolu při teplotě -16 až -24 nebo -68 až -86 °C. Jako negativní kontrola se
použije alikvotní část extraktu ze vzorku, který byl předtím testován na Clavibacter
michiganensis subsp. sepedonicus s negativním výsledkem.
Jako pozitivní kontrola
se připraví suspenze obsahující 10
5
až 106
buněk Clavibacter michiganensis subsp.
sepedonicus na ml (např. kmen NCPPB 4053 nebo PD 406) v 0,01 M fosfátového pufru
(PB) z 3-5 denní kultury (příprava viz dodatek 2). Připraví se samostatná sklíčka
s pozitivními kontrolními vzorky homologického kmene nebo jiného referenčního kmene
Clavibacter michiganensis subsp. sepedonicus suspendovaného v bramborovém extraktu
podle dodatku 2. Použití eubakteriálních oligosond značených FITC poskytuje kontrolu
procesu hybridizace, protože zbarví všechny eubakterie přítomné ve vzorku.
Test kontrolního
materiálu se provádí stejným způsobem jako u vzorků.
5.1. Fixace bramborového extraktu
5.1.1. Připraví se fixační roztok (viz dodatek 7)
5.1.2. Napipetuje se 100 µl každého vzorkového extraktu do Eppendorfovy
mikrozkumavky a odstřeďujte po dobu 8 minut na 7 000 g.
5.1.3. Odstraní se supernatant a rozpustí se peleta v 500 µl fixačního roztoku
připraveného max. 24 hodin předem. Protřepe se a inkubuje se přes noc při teplotě
4 °C.
Alternativním fixačním činidlem je 96 % etanol. Pro jeho použití se rozpustí
peleta z kroku 5.1.2. v 50 µl 0,01 M PB a 50 µl 96 % etanolu. Promíchá se protřepáním
a inkubuje se při teplotě 4 °C po dobu 30-60 minut.
5.1.4. Odstřeďuje se po dobu 8 minut na 7 000 g, odstraní se supernatant
a resuspenduje se peleta v 75 µl 0,01 M PB (viz dodatek 3).
5.1.5. Kápne se 16 µl fixované suspenze na čisté 10 okénkové sklíčko, jak
ukazuje obrázek 3, přičemž se použijí 2 různé vzorky na jedno sklíčko, a to neředěný
a zředěný 1:100 s použitím 10 µl (v 0,01 M PB). Zbývající roztok vzorku (49µl) může
být uložen při teplotě -20 °C po přidání 1 objemového množství 96 % etanolu. V případě,
že je třeba FISH metodu opakovat, odstraní se etanol odstředěním, přidá se stejné
množství 0,01 M PB a zamíchá se protřepáním.
Obrázek 3. Rozmístění na sklíčku FISH
5.1.6. Sklíčka se nechají uschnout na vzduchu (nebo sušičkou sklíček při
teplotě 37 °C) a fixují se nad plamenem.
V této fázi je možnost postup přerušit
a pokračovat v hybridizaci další den. Sklíčka by měla být skladována chráněna před
prachem a v suchu při pokojové teplotě.
5.2. Předhybridizace a hybridizace
5.2.1. Připraví se roztok lyzozymu obsahující 10 mg lyzozymu (Sigma L-6876)
v 10 ml pufru (100 mM Tris-HCl, 50 mM EDTA, pH 8,0). Tento roztok lze skladovat,
ale měl by se pouze jednou zmrazit a nechat roztát. Každý vzorek se dobře pokryje
přibližně 50 µl roztoku lyzozymu a inkubuje po dobu 10 minut při pokojové teplotě.
Potom se ponoří sklíčka do demineralizované vody, pouze jednou, a osuší filtračním
papírem.
Alternativně se přidá místo lyzozymu 50 µl proteinázy K 40-400 µg.ml-1 v
pufru (20 mM Tris-HCl, 2 mM CaCl2 , pH 7,4) do každé jamky a inkubuje se při teplotě
37 °C po dobu 30 minut.
5.2.2. Buňky se suší ve stupňovaných sériích 50 %, 80 % a 96 % etanolu pokaždé
po dobu 1 minuty. Sklíčka se nechají uschnout na vzduchu v držáku sklíček.
5.2.3. Připraví se vlhká inkubační komora zakrytím dna vzduchotěsné krabice
tkaninou nebo filtračním papírem namočeným v 1x hybmixu (dodatek 7). Krabice se přeinkubuje
v hybridizační peci při teplotě 55 °C po dobu alespoň 10 minut.
5.2.4. Připraví se hybridizační roztok (dodatek 7) s 45 µl na 1 sklíčko
a předinkubuje se po dobu 5 minut při teplotě 55 °C.
5.2.5. Sklíčka se umístí na horkou plotnu při teplotě 45 °C a do každého
ze 4 okének na sklíčku / sklíčcích se přidá 10 µl hybridizačního roztoku.
5.2.6. Každé sklíčko se přikryje 2 krycími sklíčky (24 x 24 mm) tak, aby
pod ně nevnikl vzduch. Sklíčka se umístí do předehřáté vlhké komory a hybridizuje
se 14 - 18 h v peci při teplotě 55 °C v temnu.
5.2.7. Připraví se 3 kádinky obsahující 1 l sterilní vody (Ultra pure water
= UPW), 1 l 1x hybmixu (334 ml 3x hybmix a 666 ml UPW) a 1 l 1/2 x hybmixu (167 ml
3x hybmix a 833 ml UPW). Každá z nich se přeinkubuje ve vodní lázni při teplotě 55
°C.
5.2.8. Sejmou se krycí sklíčka a podložní sklíčka se umístí do držáku sklíček.
5.2.9. Spláchne se nadbytek vzorku inkubací po dobu 15 minut v kádince s
1x hybmixem při teplotě 55 °C.
5.2.10. Držák sklíček se přemístí do promývacího roztoku 1/2 hybmix a nechá
se inkubovat dalších 15 minut.
5.2.11. Sklíčka se ponoří krátce do UPW a položí na filtrační papír. Odstraní
se nadbytečná vlhkost lehkým zakrytím povrchu filtračním papírem. Napipetuje se 5-10
µl krycího roztoku (např. Vectashield, Vecta Laboratories, CA, USA nebo podobný)
do každé jamky a celé sklíčko se zakryje velkým krycím sklíčkem (24 x 60 mm).
5.3. Hodnocení FISH testu
5.3.1. Sklíčka se prohlížejí ihned s mikroskopem vhodným pro epifluorescenční
mikroskopii se zvětšením 630x nebo 1 000x pod olejovou imerzí. S filtrem vhodným
pro fluorescein isothiokyanat (FITC) jsou eubakteriální buňky (včetně většiny gramnegativních
buněk) ve vzorku zbarveny fluorescenčně zeleně. Použitím filtru pro tetramethylrhodamin-5-isothiokyanat
se buňky Clavibacter michiganensis subsp. sepedonicus obarvené Cy3 jeví fluorescenčně
červené. Porovnává se buněčná morfologie s morfologií pozitivních kontrolních vzorků.
Buňky musí být jasně fluoreskující a zcela zbarveny. Test FISH (oddíl 9.4) musí být
zopakován, pokud je zbarvení odchylné. Prohlíží se okénka napříč dvěma průměry v
pravých úhlech a kolem obvodu. U vzorků, kde nejsou pozorovány žádné nebo málo buněk,
se pozoruje nejméně 40 polí mikroskopu.
5.3.2. Hledají se jasně fluoreskující buňky s morfologií charakteristickou
pro Clavibacter michiganensis subsp. sepedonicus v okénkách testovacích sklíček (viz
internetová stránka http://forum.europa.eu.int/Public/irc/sanco/Home/main). Intenzita
fluorescence musí odpovídat nebo být lepší než u pozitivního kontrolního kmene. Buňky,
které nejsou zcela zbarveny nebo vykazují slabou fluorescenci, se neberou v úvahu.
5.3.3. Při podezření z jakékoli kontaminace musí být test zopakován. Při
podezření z jakékoli kontaminace musí být test zopakován. To se může stát, když všechna
sklíčka ve skupině vykazují pozitivní buňky díky kontaminaci pufru nebo při zjištění
pozitivních buněk (mimo okénka sklíček) na povrchu sklíček.
5.3.4. Existuje několik problémů podstatných pro přesnost FISH testu. V peletách
z pupkové části bramboru a částí stonku se mohou vyskytnout doprovodné populace fluoreskujících
buněk s atypickou morfologií a křížově reagující saprofytické bakterie s velikostí
a morfologií podobnou Clavibacter michiganensis subsp. sepedonicus, ačkoli mnohem
méně často než u IF testu.
5.3.5. V úvahu se berou pouze fluoreskující buňky s typickou velikostí a
morfologií, viz 5.3.2.
5.3.6. Interpretace výsledku testu FISH:
a) Výsledky FISH testu jsou platné, pokud jsou při použití FITC filtru
jasně zeleně fluoreskující buňky s velikostí a morfologií typickou pro Clavibacter
michiganensis subsp. sepedonicus a při použití rhodaminového filtru jasně červeně
fluoreskující buňky pozorovány ve všech pozitivních kontrolách a nejsou pozorovány
v žádných negativních kontrolách. Pokud jsou přítomné jasně fluoreskující buňky s
typickou morfologií, odhadne se průměrný počet typických buněk v 1 mikroskopickém
poli a vypočítá počet typických buněk v 1 ml resuspendované pelety (dodatek 4). Vzorky,
které obsahují alespoň 5 x 10
3
typických buněk na 1 ml resuspendované pelety, se
považují za pravděpodobně infikované Clavibacter michiganensis subsp. sepedonicus.
Nutné je další testování. Vzorky, které obsahují méně než 5 x 103
typických buněk
na 1 ml resuspendované pelety, se považují za negativní.b) Výsledek FISH testu je negativní, pokud při použití rhodaminového filtru
nejsou pozorovány jasně červeně fluoreskující buňky s velikostí a morfologií typickou
pro Clavibacter michiganensis subsp. sepedonicus, jestliže jsou tyto typické jasně
červeně fluoreskující buňky při použití rhodaminového filtru pozorovány v pozitivních
kontrolách.
6. PCR test
PRINCIP
Použije-li se PCR test jako hlavní screeningový test a je pozitivní,
musí být jako druhý povinný screeningový test proveden IF test. Pokud se PCR test
používá jako druhý screeningový test a je pozitivní, je pro dokončení diagnózy nutné
další testování podle postupového diagramu.
Využití této metody v celém rozsahu jako
hlavního screeningového testu se doporučuje jen tehdy, je-li požadována specializovaná
expertíza.
Poznámka:
Předběžné testování touto metodou by mělo umožnit reprodukovatelné
zjištění 10
3
až 104
buněk Clavibacter michiganensis subsp. sepedonicus na 1 ml přidaných
do vzorku extraktů, které byly předtím testovány s negativním výsledkem. Pro dosažení
maximální citlivosti a přesnosti ve všech laboratořích mohou být vyžadovány optimalizační
pokusy. Používají se validovaná PCR činidla a protokoly. Přednostně se používá metoda
s interní kontrolou.
Je třeba použít vhodná bezpečnostní opatření, aby se zabránilo
kontaminaci vzorku cílovou DNA. PCR test by měli provádět zkušení laboranti v laboratořích
specializovaných na molekulární biologii, aby se minimalizovala možnost kontaminace
cílovou DNA.
S negativními kontrolami (u průběhu extrakce DNA a PCR) by se mělo vždy
zacházet jako s konečnými vzorky, aby bylo jasné, jestli došlo k přenosu DNA.
PCR test by měl zahrnovat následující negativní kontroly:
- extraktu ze
vzorku, který byl předtím testován na Clavibacter michiganensis subsp. sepedonicus
s negativním výsledkem,
- kontroly pufru používaného pro extrakci bakterie a DNA
ze vzorku,
- reakční směs PCR.
Měly by být zahrnuty následující pozitivní kontroly:
- alikvotní části
resuspendovaných pelet, k nimž byl přidán Clavibacter michiganensis subsp. sepedonicus
(příprava viz dodatek 2),
- suspenze 106 buněk na 1 ml Clavibacter michiganensis
subsp. sepedonicus ve vodě z virulentního izolátu (např. NCPPB 2140 nebo NCPPB 4053),
- pokud možno použít při provádění PCR testu také DNA extrahovanou z pozitivních
kontrolních vzorků.
Aby se zabránilo možné kontaminaci, připraví se pozitivní kontroly v odděleném
prostředí od vzorků, které budou testovány.
Extrakty ze vzorků by měly být pokud možno bez zeminy. V případě použití
PCR testu je potřeba připravit extrakty z umytých brambor.
6.1 Metody purifikace DNA
Použijí se výše popsané pozitivní a negativní kontrolní vzorky.
Připraví se kontrolní materiál stejným způsobem jako vzorky.
K purifikaci cílové DNA z komplexních substrátů vzorků jsou k dispozici
různé metody odstraňující inhibitory PCR a jiných enzymatických reakcí a koncentrující
cílovou DNA v extraktu vzorku.
Následující metoda byla optimalizována pro použití s validovanou metodou
PCR, uvedenou v dodatku 6.
6.1. a) Metoda podle Pastrika (2000)
1. Napipetuje se 220 µl lýzového pufru (100 mM NaCl, 10 mM Tris-HCl [pH
8,0], 1 mM EDTA [pH 8,0]) do mikrozkumavky o objemu 1,5 ml.
2. Přidá se 100 µl extraktu
ze vzorku a umístí se do termobloku nebo vodní lázně o teplotě 95 °C na dobu 10 minut.
3. Mikrozkumavka se vloží na 5 minut do ledu.
4. Přidá se 80 µl zásobního roztoku
lyzozymu (50 mg lyzozymu na 1 ml v 10 mM Tris HCl, pH 8,0) a inkubuje se při teplotě
37 °C po dobu 30 minut.
5. Přidá se 220 µl Easy DNA(R) roztok A (Invitrogen), dobře
se promíchá třepáním a inkubuje se při teplotě 65 °C po dobu 30 minut.
6. Přidá se
100 µl Easy DNA (R) roztok B (Invitrogen)a silně se promíchá třepáním, až usazenina
sama poteče do mikrozkumavky a vzorek začne být stejnoměrně viskózní.
7. Přidá se
500 µl chloroformu a míchá se třepáním, až se viskozita sníží a směs se stane homogenní.
8. Odstřeďuje se na 15 000 g po dobu 20 minut při 4 °C pro oddělení fází a vytvoření
mezifáze.
9. Přenese se horní fáze do čisté mikrozkumavky.
10. Přidá se 1 ml 100
% etanolu (-20 °C), krátce se promíchá třepáním a inkubuje se na ledu po dobu 10
minut.
11. Odstředí se na 15 000 g po dobu 20 minut při 4 °C a odstraní se z pelety
etanol.
12. Přidá se 500 µl 80 % etanolu (-20 °C) a promíchá převracením mikrozkumavky.
13. Odstřeďuje se na 15 000 g po dobu 10 minut při 4°C, peleta se zachová a odstraní
etanol.
14. Peleta se nechá uschnout na vzduchu nebo v DNA speed vac.
15. Peleta
se resuspendujte v 100 µl sterilní vody (UPW) a nechá stát nejméně 20 minut při pokojové
teplotě.
16. Skladuje se při teplotě -20 °C až do upotřebení při PCR.
17. Jakákoliv
bílá usazenina se odstraní odstředěním a pro PCR se použije 5 µl supernatantu obsahujícího
DNA.
6.1.b) Jiné metody
Jiné metody extrakce DNA by se mohly použít, pokud by se prokázalo, že jsou
při purifikaci DNA z kontrolních vzorků obsahujících 10
3
až 104
patogenních buněk
na 1 ml stejně efektivní.6.2 PCR
6.2.1. Připraví se testované vzorky a kontroly pro PCR podle validovaného
protokolu (dodatek 6). Připraví se jeden desetinný roztok vzorku DNA (1:10 ve sterilní
vodě).
6.2.2. Připraví se příslušná reakční směs pro PCR v prostředí, kde nehrozí
kontaminace, podle zveřejněného protokolu (dodatek 6). Validovaný protokol PCR je
multiplexová reakce, která zahrnuje také interní kontrolu PCR.
6.2.3. Do sterilních PCR mikrozkumavek se přidá 5 µl extraktu DNA na 25
µl PCR reakce.
6.2.4. Zahrne se negativní kontrolní vzorek obsahující pouze reakční směs
PCR a přidá se stejná sterilní voda UPW, která byla použita do směsi PCR namísto
vzorku.
6.2.5. PCR mikrozkumavky se umístí do stejného termocykleru, který byl použit
při počátečním testování a provede se vhodně optimalizovaný program PCR (dodatek
6)
6.3. Analýza produktu PCR
6.3.1. Rozdělí se amplikony PCR elektroforézou v agarózovém gelu. Nanese
se nejméně 12 µl amplifikované reakční směsi DNA z každého vzorku smíchané s 3 µl
nanášecího pufru (dodatek 6) do 2,0 % (w/v) agarózového gelu v Trisacetát-EDTA (TAE)
pufru (dodatek 6) při 5-8 V na cm. Použije se vhodný marker DNA, např. 100 bp ladder.
6.3.2. Detekují se proužky DNA barvením v ethidium bromidu (0,5 mg na l)
po dobu 30-45 minut, za použití vhodných bezpečnostních opatření pro zacházení s
tímto mutagenem.
6.3.3. V obarveném a UV (krátké vlnové délky, např. 302 nm) prosvíceném
gelu se hledají amplifikované produkty PCR o očekávané velikosti a výsledek se zdokumentuje.
6.3.4. U všech nových nálezů se zkontroluje pravost amplikonu PCR provedením
restrikční enzymové analýzy ve zbývajícím vzorku amplifikované DNA inkubací při optimální
teplotě a době s vhodným restrikčním enzymem a pufrem (viz dodatek 6). Rozdělí se
naštěpené fragmenty elektroforézou v agarózovém gelu a pozoruje se charakteristický
vzor restrikčního fragmentu pod UV prosvícením po obarvení ethidiumbromidem a porovnává
se s neštěpenou a štěpenou pozitivní kontrolou.
Interpretace výsledku PCR testu: PCR test je negativní, pokud amplikon typický
pro
Clavibacter michiganensis subsp. sepedonicus očekávané velikosti nebyl zjištěn
v daném vzorku, ale byl zjištěn ve všech pozitivních kontrolních vzorcích (v případě
vícenásobné PCR s rostlinnými interními kontrolními primery: druhý produkt PCR očekávané
velikosti musí být amplifikován s daným vzorkem).
PCR test je pozitivní, pokud byl zjištěn amplikon typický pro Clavibacter
michiganensis subsp. sepedonicus očekávané velikosti a vzoru (pokud se požaduje),
za předpokladu, že nebyl amplifikován žádným z negativních kontrolních vzorků. Spolehlivého
potvrzení pozitivního výsledku lze dosáhnout také opakováním testu s druhou sadou
PCR primerů (oddíl 9.3).
Poznámka:
Lze mít podezření na inhibici PCR, pokud byl očekávaný amplikon
získán z pozitivního kontrolního vzorku obsahujícího Clavibacter michiganensis subsp.
sepedonicus ve vodě, ale z pozitivní kontroly s Clavibacter michiganensis subsp.
sepedonicus v bramborovém extraktu byly negativní. Ve vícenásobných protokolech PCR
s interními kontrolami PCR i inhibici reakce došlo, pokud nebyl získán žádný z obou
amplikonů.
Lze mít podezření na kontaminaci, pokud byl očekávaný amplikon získán z
jedné nebo více negativních zkoušek.
7. Test na lilku vejcoplodém
Poznámka:
Předběžné testování touto metodou by mělo umožnit reprodukovatelné
zjištění 10
3
až 104
jednotek Clavibacter michiganensis subsp. sepedonicus tvořících
kolonie na 1 ml přidaný k extraktům ze vzorku, které byly testovány s negativním
(příprava viz. dodatek 2) Nejvyšší citlivost zjištění lze očekávat při použití čerstvě
připraveného extraktu ze vzorku a optimálních růstových podmínkách. Metodu však lze
úspěšně použít i na extrakty, které byly uchovávány v glycerolu při teplotě -68 až
-86 oC.
Některé odrůdy lilku vejcoplodého poskytují vynikající selektivní obohacující
médium pro růst Clavibacter michiganensis subsp. sepedonicus dokonce v případě nepřítomnosti
příznaků a poskytují také vynikající základní konfirmační test.
Pro omezení nebezpečí falešně negativních výsledků by měly být vytvořeny
optimální růstové podmínky.
Podrobnosti o pěstování jsou uvedeny v Dodatku 8.
7.1. Peleta podle 3.1.5. se rozdělí mezi rostliny lilku jednou z níže uvedených
metod (7.2, 7.3, 7.4). Používají se pouze rostliny ve fázi 2-3 listů do úplného rozvinutí
třetího pravého listu. Aby se zajistilo úplné využití resuspendované pelety i efektivní
inokulaci, bude pro níže uvedené postupy potřeba 15-25 lilků na vzorek.
7.2. Inokulace zářezem I
7.2.1. Každý květináč se horizontálně podepře (pro květináče o průměru 10
cm je vhodný blok pěnového polystyrénu s vyhloubenou částí o rozměrech 5 cm hloubky,
10 cm šířky a 15 cm délky). Pro každý testovaný vzorek se mezi stonek a blok umístí
proužek sterilní hliníkové fólie. Rostlinu je možno fixovat na místě gumovým páskem
kolem bloku.
7.2.2. Pomocí skalpelu se provede mezi děložními lístky a prvním pravým
listem podélný nebo mírně úhlopříčný řez dlouhý 0,5 až 1,0 cm a hluboký přibližně
jako tři čtvrtiny průměru stonku.
7.2.3. Zářez se přidrží otevřený pomocí špičky čepele skalpelu a nanese
se do něj inokulum štětečkem na oční linky nebo jemným malířským štětečkem namočeným
do pelety. Zbytek pelety se rozdělí mezi všechny testovací rostliny lilku.
7.2.4. Řez se překryje sterilní vazelínou aplikovanou injekční stříkačkou
o objemu 2 ml.
7.3 Inokulace zářezem II
7.3.1. Na stonek rostliny držené dvěma prsty se napipetuje mezi děložní
lístky a první pravý list kapka (přibližně 5 až 10 µl) suspendované pelety.
7.3.2. Pomocí sterilního skalpelu se udělá sešikmený zářez (v úhlu přibližně
5°), dlouhý 1,0 cm a hluboký přibližně jako 2/3 tloušťky stonku, přičemž s řezem
se začne v místě kapky suspendované pelety.
7.3.3. Řez se zakryje sterilní vazelínou z injekční stříkačky.
7.4. Inokulace injekční stříkačkou
7.4.1 Pro snížení vnitřního napětí buněk (turgoru) se rostliny lilku den
před inokulací nezalévají.
7.4.2 Stonky lilku vejcoplodého se inokulují těsně nad děložními lístky
pomocí injekční stříkačky s podkožní jehlou (ne méně než 23 G). Peleta se rozdělí
mezi testovací rostliny lilku vejcoplodého.
7.5. Jako pozitivní kontrola se inokuluje 5 rostlin stejnou inokulační metodou
(7.2, 7.3 nebo 7.4) vodní suspenzí 10
5
až 106
buněk na 1 ml známé kultury původce
kroužkovitosti a kde je to možné i pletivem z přirozeně infikovaných hlíz bramboru.7.6. Jako negativní kontrola se inokuluje 5 rostlin sterilním 0,05 M PBS
stejnou inokulační metodou (7.2, 7.3 nebo 7.4).
7.7. Po inokulaci se rostliny nechají inkubovat v karanténě ve vhodných podmínkách
(dodatek 8) po dobu až 4 týdnů při teplotě 18-24°C. Rostliny se inkubují za dostatečného
osvětlení a vysoké vlhkosti (70-80%) a zalévají se tak, aby nedošlo k nasávání vody
nebo vadnutí kvůli nedostatku vody. Buňky Clavibacter michiganensis subsp. sepedonicus
odumírají při teplotách nad 30°C a optimální teplota je 21°C. Aby se zamezilo kontaminaci,
inkubují se rostliny pro pozitivní a negativní kontroly ve skleníku nebo růstové
komoře na jasně oddělených policích, anebo při nedostatku místa se zajistí přísné
oddělení mezi zacházení s nimi. Pokud musí být rostliny pro různé vzorky inkubovány
blízko sebe, oddělí se vhodnými přepážkami. Při hnojení, zalévání, kontrole a jiné
manipulaci se věnuje maximální pozornost tomu, aby nedošlo ke kontaminaci. Je nezbytné,
aby skleníky i růstové komory byly chráněny před veškerým hmyzem, protože by mohl
přenášet bakterie z jednoho vzorku na druhý.
7.8. Pravidelně po osmi dnech se spočítají rostliny vykazující příznaky.
Původce kroužkovitosti působí u lilku vejcoplodého vadnutí listů, které může začít
jako okrajová nebo mezižilková ochablost (ztráta turgoru). Zvadlé pletivo může zprvu
být tmavozelené nebo strakaté, ale před znekrotizováním zesvětlí. Povadlá místa mezi
žilnatinou mívají často mastně vodnatý vzhled. Nekrotická pletiva mívají často jasně
žlutý okraj. Rostliny vždy neodumírají; čím déle trvá období do objevení se příznaků,
tím je větší naděje na přežití. Rostliny mohou infekci odrůst. Mladé rostliny lilku
vejcoplodého jsou mnohem citlivější vůči nízkým koncentracím původce kroužkovitosti
než starší rostliny, proto je nezbytné používat rostliny ve fázi tří listů a nebo
krátce před ní. Vadnutí mohou také způsobovat populace jiných bakterií nebo hub přítomných
v peletě z pletiv hlízy. Patří k nim Erwinia carotovora subsp. carotovora a E. carotovora
subsp. atroseptica, Phoma exigua var. foveata, jakož i velké koncentrace saprofytických
bakterií. Tyto případy vadnutí lze odlišit od případů způsobených původcem kroužkovitosti,
jelikož rychle vadnou celé listy nebo celé rostliny. Může se také připravit Gramovo
barvení: tento test rozliší Clavibacter michiganensis subsp. sepedonicus od Erwinia
spp
7.9. Jakmile se na lilku vejcoplodém objeví příznaky, měla by být provedena
izolace za použití částí pletiva zvadlých listů nebo stonků rostlin. Povrch listů
a stonků lilku vejcoplodého se vydezinfikuje otřením 70 % etanolem. Provede se test
IF nebo PCR na šťávě z lilku a izoluje se na vhodném (selektivním) médiu (viz oddíl
8). Může se také připravit Gramovo barvení (dodatek 9). Čisté kultury podezřelé z
přítomnosti Clavibacter michiganensis subsp. sepedonicus se identifikují a potvrdí
se patogenita (viz oddíl 9 a 10).
7.10. Za určitých okolností, zejména pokud nejsou růstové podmínky optimální,
se může stát, že Clavibacter michiganensis subsp. sepedonicus zůstává v rostlinách
lilku vejcoplodého jako latentní infekce dokonce i po uplynutí inkubační doby až
4 týdnů. Pokud nejsou po 4 týdnech pozorovány žádné příznaky, provede se test IF/PCR
na složeném vzorku částí stonků o délce 1 cm z každé testované rostliny odebraných
nad místem inokulace. Pokud je test pozitivní, měla by být provedena izolace na vhodných
(selektivních) médiích postupem podle oddílu 8. Čisté kultury podezřelé z přítomnosti
Clavibacter michiganensis subsp. sepedonicus se identifikují a potvrdí se patogenita
(viz oddíl 9 a 10).
8. Izolace Clavibacter michiganensis subsp. sepedonicus
Diagnóza může být potvrzena pouze tehdy, je-li původce kroužkovitosti izolován
a takto identifikován. Ačkoliv je původce kroužkovitosti náročným organismem, je
možno ho izolovat z pletiv projevujících příznaky napadení.
Mohou jej však přerůst rychle rostoucí saprofytické bakterie, a proto se
nedoporučuje provádět izolaci přímo z pelety pletiva hlízy (3.1.5) nebo stonku (3.2).
Přímá izolace Clavibacter michiganensis subsp. sepedonicus je možná za použití selektivního
média a vhodného ředění resuspendované pelety z pupkové části hlízy nebo ze stonků
rostliny.
Izolace se provádí ze všech hlíz bramboru nebo částí stonku a z rostlin
lilku vejcoplodého, které nevykazují příznaky napadení, ale u nichž byl pozitivní
výsledek v testu IF/PCR složených vzorků (viz oddíl 7.9). Pokud je třeba provést
maceraci stonků lilku, měla by být provedena podle oddílu 3.1.2.
Jako pozitivní kontroly se připraví desetinná ředění suspenze 106 cfu na
ml Clavibacter michiganensis subsp. sepedonicus (např. NCPPB 4053 nebo PD 406). Aby
se vyloučilo nebezpečí kontaminace, připraví se pozitivní kontroly odděleně od vzorků,
které mají být testovány.
Pro každou nově připravenou dávku selektivního média by se měla před použitím
k testování obvyklých vzorků přezkoušet jeho vhodnost pro růst patogenu.
Kontrolní materiál se testuje stejným způsobem jako vzorky.
8.1. Roztěr na selektivní médium
8.1.1. Ze 100 µl alikvotní části ze vzorku resuspendované bramborové pelety
nebo šťávy z lilku vejcoplodého se připraví desetinásobné ředění v peletovém pufru
(dodatek 3).
8.1.2. Izolace z neředěné bramborové pelety se obvykle nepodaří kvůli náročným
růstovým podmínkám Clavibacter michiganensis subsp. sepedonicus a konkurenci saprofytů.
Vzhledem k tomu, že bakterie je obvykle v infikovaných pletivech přítomná ve vysokých
koncentracích, lze saprofyty obvykle vypláchnout ředěním, zatímco patogen zůstává.
Proto se doporučuje rozetřít 100 µl z každého vzorku, v ředění1/100 až 1/10 000 na
MTNA médium nebo NCP- 88 médium (dodatek 5), za použití pomůcek určených k roztěrům
(hokejek) a techniky roztěrů.
8.1.3. Desky se inkubují v temnu při teplotě 21-23°C.
8.1.4. Počáteční kontrola misek zahrnuje porovnání s kontrolními miskami
a počítání všech kolonií podobných Clavibacter michiganensis subsp. sepedonicus se
provádějí po třech dnech, s dalším počítáním po 5,7 a 10 dnech.
8.2. Čištění podezřelých kolonií
Poznámka:
Subkultury kolonií podobných Clavibacter michiganensis subsp.
sepedonicus pro inokulaci lilku vejcoplodého a/nebo následnou identifikaci by se
měly pěstovat na YGM médiu; inokulace a identifikace by se měly provést dříve, než
jsou média příliš přerostlá, tj. nejlépe po 3-5 dnech.
8.2.1. Kolonie podobné Clavibacter michiganensis subsp. sepedonicus se rozetřou
na jedno z následujících médií (složení uvedena v dodatku 5):
- živný dextrózový
agar (pouze pro subkultury),
- kvasničný pepton glukosový agar,
- agar z kvasničného
extraktu s minerálními solemi.
Inkubace probíhá při teplotě 21-24 °C po dobu maximálně
10 dní.
Původce kroužkovitosti roste pomalu a obvykle vytváří kolonie velikosti špendlíkové
hlavičky, vyklenuté, smetanově zbarvené.
8.2.2. Opětovně se provede roztěr pro zaručení čistoty. U subkultur se rychlost
růstu zlepšuje. Typické kolonie jsou smetanově bílé nebo barvy slonoviny, někdy žluté,
okrouhlé, hladké, vyvýšené, konvexně vyklenuté, slizovitě tekuté, s rovnými okraji
a obvykle mají v průměru 1 - 3 mm.
Jednoduché Gramovo barvení (dodatek 9) může pomoci
vybrat kolonie pro další testování.
8.2.3 Podezřelé kultury se identifikují (viz oddíl 9) a provede se zkouška
patogenity (viz oddíl 10).
9. Identifikace
Čisté kultury pravděpodobné izolované kultury Clavibacter michiganensis
subsp. sepedonicus se identifikují za použití nejméně dvou následujících testů založených
na různých biologických principech. V případě potřeby se zahrne pro každý provedený
test známý referenční kmen.
9.1. Nutriční a enzymatické identifikační testy
Zjišťují se následující
fenotypické vlastnosti. Veškerá média by se měla inkubovat při 21 °C a po šesti dnech
by měla být vyhodnocena. Pokud nedošlo k žádnému růstu, inkubuje se po dobu nejvýše
20 dní.
Všechny testy musí zahrnovat kontrolu se známým kmenem Clavibacter michiganensis
subsp. sepedonicus. Nutriční a fyziologické testy se musí provádět za použití inokula
ze subkultur živného agaru. Morfologická srovnání se musí provádět z kultur z živného
dextrózového agaru.
Test Očekávaný výsledek
Oxidačně-fermentační (O/F) inertní nebo slabě
oxidační
Aktivita oxidázy -
Aktivita katalázy +
Redukce nitrátů -
Aktivita ureázy -
Tvorba H2S -
Tvorba indolu -
Využívání citrátu -
Hydrolýza škrobu - nebo slabá
Růst při 37 °C -
Růst v 7% NaCl -
Hydrolýza želatiny -
Hydrolýza eskulinu +
Tvorba kyseliny z:
- glycerolu -
- laktózy - nebo slabá
- rhamnózy -
- salicinu -
Gramovo barvení (dodatek 9)
9.2. IF test
a) Připraví se suspenze přibližně 10
6
buněk/ml v pufru na IF (dodatek 3). b) Připraví se série dvojnásobného ředění vhodného antiséra.
c) Provede se IF postup (oddíl 4).
d) Pozitivního výsledku IF testu je dosaženo, jestliže IF titr kultury
odpovídá titru pozitivní kontroly.
9.3. PCR test
a) Připraví se suspenze přibližně 106 buněk na ml ve sterilní vodě (UPW)
b) Zahřívá se 100 µl suspenze buněk v uzavřených mikrozkumavkách v ohřívacím
bloku nebo vřící vodní lázni při teplotě 100 °C po dobu 4 minut. V případě potřeby
se může lýza buněk podpořit přidáním čerstvě připraveného NaOH do konečné koncentrace
0,05 M. Vzorky lze potom uložit při teplotě -16 až -24 °C až do použití.
c) Pro amplifikaci Clavibacter michiganensis subsp. sepedonicus specifických
amplikonů se použijí vhodné postupy PCR (např. Pastrik, 2000; viz dodatek 4; Li a
de Boer, 1995; Mills a kol., 1997; Pastrik a Rainey, 1999; Schaad a kol., 1999).
d) Identifikace Clavibacter michiganensis subsp. sepedonicus je pozitivní,
pokud jsou amplikony PCR stejné velikosti a mají stejnou mnohotvárnost délky fragmentu
jako pozitivní kontrolní kmen.
9.4. FISH test
a) Připraví se suspenze přibližně 106 buněk na ml v UPW.
b) Provede se postup FISH (oddíl 5)
c) Test FISH je pozitivní, jsou-li dosaženy stejné reakce kultury a pozitivní
kontroly.
9.5. Analýza mastných kyselin (FAP)
a) Kultura se pěstuje na tryptikázo-sójovém agaru (Oxoid) po dobu 72 hodin
při teplotě 21 °C (+/- 1°C).
b) Použije se vhodný postup FAP (Janse, 1991; Stead, 1992).
c) Test FAP je pozitivní, pokud je profil podezřelé kultury identický
s profilem pozitivní kontroly. Přítomnost charakteristických mastných kyselin 15:1
Anteiso A, 15:0 Iso, 15:0 Anteiso, 16:0 Iso, 16:0 a 17:0 Anteiso jasně svědčí o přítomnosti
Clavibacter michiganensis subsp. sepedonicus. Jiné rody jako Curtobacterium, Arthrobacter
a Micrococcus také obsahují některé z těchto kyselin, ale 15:1 Anteiso A je pro tyto
bakterie neobvyklá kyselina, která se však vyskytuje ve všech Clavibacter spp. v
rozmezí 1-5 %. U Clavibacter michiganensis subsp. sepedonicus se hodnota obvykle
pohybuje kolem 5 %.
9.6. BOX-PCR
a) Připraví se suspenze přibližně 106 buněk na ml v UPW.
b) Provede se test postupem podle Smith a kol., 2001.
10. Test patogenity
Pro konečné potvrzení původce kroužkovitosti a pro
stanovení virulence kultur identifikovaných jako Clavibacter michiganensis subsp.
sepedonicus musí být provedena zkouška patogenity.
10.1. Připraví se inokulum přibližně 10
6
buněk na ml z 3- denních kultur
testované izolované látky a vhodného pozitivního kmene Clavibacter michiganensis
subsp. sepedonicus.10.2. Naočkuje se 5-10 stonků mladých semenáčků lilku vejcoplodého ve fázi
3 pravých listů.
10.3. Inkubuje se při teplotě 18-24 °C při dostatečném světle a vysoké relativní
vlhkosti s přiměřeným zaléváním, aby nedošlo k přemokření nebo vyschnutí. U čistých
kultur by mělo během 2 týdnů nastat typické vadnutí, avšak rostliny, které po uplynutí
této doby nevykazují žádné příznaky infekce, by se měly inkubovat až 3 týdny při
teplotách příznivých pro růst lilku vejcoplodého, ale nepřesahujících 25 °C. Jestliže
se po 3 týdnech příznaky infekce nevyskytují, nemůže být kultura považována za patogenní
formu Clavibacter michiganensis subsp. sepedonicus.
10.4. Izolace se provádí z rostlin s příznaky infekce odstraněním části
stonku 2 cm nad místem inokulace. Pletiva se rozdrtí a suspendují v malém množství
sterilní destilované vody nebo 50 mM fosfátového pufru. Izoluje se ze suspenze rozetřením
nebo nanesením na MTNA a YPGA, inkubuje se po dobu 3-5 dní při teplotě 21-23°C a
sleduje se vznik kolonií typických pro Clavibacter michiganensis subsp. sepedonicus.
Dodatek 1
Laboratoře podílející se na optimalizaci a validaci protokolů
------------------------------------------------------ -------------------------------------------
Laboratoř (1) Místo Země
------------------------------------------------------ -------------------------------------------
Agentur für Gesundheit und Vídeň a Linec Rakousko
Ernährungssicherheit
Departement Gewasbescherming Merelbeke Belgie
Plantedirektoratet Lyngby Dánsko
Central Science Laboratory York Anglie
Scottish Agricultural Science Agency Edinburgh Skotsko
Laboratoire National de la Protection Angers Francie
Végétaux, Unité de Bactériologie
Laboratoire National de la Protection Le Rheu Francie
Végétaux, Station de Quarantaine de la Pomme
de Terre
Biologische Bundesanstalt Kleinmachnow Německo
Pflanzenschutzamt Hannover Hannover Německo
State Laboratory Dublin Irsko
Plantenziektenkundige Dienst Wageningen Nizozemsko
Norwegian Crop Research Institute, Plant Aas Norsko
Protection Centre
Direcçăo-General de Protecçăo das Culturas Lisabon Portugalsko
Nacionalni institut za biologijo Ljubljana Slovinsko
Centro de Diagnóstico de Aldearrubia Salamanca Španělsko
------------------------------------------------------ -------------------------------------------
(1) Kontaktní osoby: viz internetová stránka http://forum.europa.eu.int/Public/irc/sanco/Home/main
Dodatek 2
Příprava pozitivních a negativních kontrol pro vyšetření výkrojků testy
PCR/IF a FISH
Vypěstuje se 72 hodinová kultura virulentního kmene C. m. subsp. sepedonicus
(NCPPB 4053 nebo PD 406) na základním médiu MTNA a suspenduje se v 10 mMfosfátového
pufru pro získání hustoty přibližně 1 až 2 x 10
8
buněk schopných tvorby kolonií/ml.
Toho se obvykle dosáhne prostřednictvím slabě zakalené suspenze odpovídající optické
hustotě 0,20 - 600 nm. Výkrojky pletiva se odeberou z pupkových konců 200 hlíz odebraných
z produkce odrůdy s bílou slupkou, u které je jisté, že je prosta C. m. subsp. sepedonicus.Pupkové výkrojky se zpracují obvyklým způsobem a resuspenduje se peleta v
10 ml.
Připraví se 10 sterilních mikrozkumavek o objemu 1,5 ml s 900 mikrol resuspendované
pelety.
Přenese se 100 ěl suspenze C. m. subsp. sepedonicus do první mikrozkumavky.
Nechá se protřepat.
V následujících pěti mikrozkumavkách se provedou desetinná ředění.
Šest kontaminovaných mikrozkumavek se použije jako pozitivní kontrola. Čtyři
nekontaminované mikrozkumavky se použijí jako negativní kontroly. Mikrozkumavky se
opatří štítkem.
Připraví se alikvotní části z 100 mikrol ve sterilních mikrozkumavkách o
objemu 1,5 ml, čímž se získá 9 kopií každého kontrolního vzorku. Skladování probíhá
při teplotě - 16 až - 24 °C až do doby použití.
Přítomnost a množství C. m. subsp. sepedonicus v kontrolních vzorcích by
měla být nejprve potvrzena prostřednictvím imunofluorescence.
Pro PCR test se provede extrakce DNA z pozitivních a negativních kontrolních
vzorků pro každou sérii zkušebních vzorků.
Pro IF a FISH testy se provedou kvantitativní rozbory pozitivních a negativních
kontrolních vzorků pro každou sérii zkušebních vzorků.
Při kvantitativních rozborech IF, FISH a PCR musí být C. m. subsp. sepedonicus
zjištěn v nejméně v 10
6
a 104
buněk/ml pozitivních kontrol a nesmí být zjištěn v
žádné z negativních kontrol.Dodatek 3
Pufry pro testovací postupy
OBECNĚ: Neotevřené sterilizované pufry lze skladovat po dobu až jednoho roku.
1. Pufry pro extrakci
1.1.Extrakční pufr (50 mM fosfátový pufr, pH 7,0)
Tento pufr se používá k extrakci bakterie z rostlinných tkání homogenizací
nebo protřepáním.
Na2HPO4(bezvodý) 4,26 g KH2PO42,72 g Destilovaná voda 1,00 l
Složky se rozpustí, zkontroluje se pH a provede se sterilizace v autoklávu
při 121 °C po dobu 15 min.
Užitečné mohou být následující složky:
Účel Množství (na
litr)
Lubrolové vločky Protisrážlivý prostředek (*) 0,5 g
DC silikonový odpěňovač Odpěňovací činidlo (*) 1,0 ml
Tetrasodiumpyrofosfát Antioxidační činidlo 1,0 g
Polyvinylpyrrolidon-40 000 (PVP - 40) Vázání inhibitorů PCR 50 g
-------------------------------------------
(*) pro použití při extrakci homogenizací
1.2.Peletový pufr (10 mM fosfátový pufr, pH 7,2)
Tento pufr se používá pro resuspenzi a ředění extraktů z výkrojků z pupkových
částí bramborových hlíz poté, co byly odstřeďováním koncentrovány do pelety.
Na2HPO4.12H2O 2,7 g NaH2PO4. 2H2O 0,4 g Destilovaná voda 1,00 l
Složky se rozpustí, zkontroluje se pH a provede se sterilizace v autoklávu
při 121 °C po dobu 15 min.
2. Pufry pro IF test
2.1. Pufry pro IF (10 mM fosfátový pufr ve fyziologickém roztoku (PBS), pH
7,2)
Tento pufr se používá k ředění protilátek. Na2HPO4.12H2O 2,7 g NaH2PO4. 2H2O 0,4 g NaCl 8,0 g Destilovaná voda 1,00 l
Složky se rozpustí, zkontroluje se pH a provede se sterilizace v autoklávu
při 121 °C po dobu 15 min
2.2. IF-pufr-Tween
Tento pufr se používá k mytí sklíček. Přidá se 0,1 % Tween 20 k pufru pro
IF.
2.3. Fosfátový pufr v glycerolu, pH 7,6
Tento pufr se používá jako krycí roztok na okénka sklíček na IF testy k zvýšení
fluorescence.
Na2HPO4.12H2O 3,2 g NaH2PO4. 2H2O 0.15 g Glycerol 50 ml Destilovaná voda 100 ml
Krycí roztoky jsou komerčně dostupné, např. Vectashield (R) (Vector Laboratories)
nebo Citifluor® (Leica).
Dodatek 4
Stanovení koncentrace IF a FISH pozitivních buněk
1. Vypočítá se průměrný počet typických fluoreskujících buněk v jednom pozorovacím
poli (c).
2. Vypočítá se počet typických fluoreskujících buněk v okénku mikroskopického
sklíčka (C).
C = c x S/s,
kde S = plocha jednoho pole na sklíčku s více jamkami a
s
= plocha pole objektivu.
s = đi2/4G2K 2,
kde i = koeficient pole (v rozmezí od 8 - 24 podle typu
okuláru),
K = tubusový koeficient (1 nebo 1,25),
G = zvětšení objektivu (100 x, 40
x atd.).
3. Vypočítá se počet charakteristických fluoreskujících buněk na 1 ml resuspendované
pelety (N).
N = C x 1 000/y x F,
kde y = objem resuspendované pelety v každém okénku
a
F = zřeďovací faktor resuspendované pelety
Dodatek 5
Média pro izolaci a kultivaci C. m. subsp. sepedonicus
1. Obecná růstová média
Živný agar (Nutrient agar = NA)
Živný agar (Difco) 23 g Destilovaná voda 1,00 l
Složky se rozpustí a provede se sterilizace v autoklávu při 121 °C po dobu
15 min.
Živný dextrózový agar (Nutrient dextrose agar = NDA)
Difco bakto živný agar obsahující 1 % D(+) glukózy (monohydrátu). Provede
se sterilizace v autoklávu při 121 C po dobu 20 min.
Kvasnično-pepton-glukózový agar (Yeast peptone glucose agar = YPGA)
Kvasnicový extrakt (Difco) 5,0 g Baktopepton (Difco) 5,0 g D(+) glukóza (monohydrát) 10,0 g Baktoagar (Difco) 15,0 g Destilovaná voda 1,00 l
Složky se rozpustí a provede se sterilizace v autoklávu při 121 °C po dobu
15 min.
Médium s kvasnicovým extraktem a minerálními solemi (Yeast extract mineral
salts medium = YGM)
Kvasnicový extrakt 2,0 g (Difco) D(+) glukóza 2,5 g (monohydrát) K2HPO40,25 g KH2PO40,25 g MgSO4. 7H2O 0,1 g MnSO4. H2O 0,015 g NaCl 0,05 g FeSO4. 7H2O 0,005 g Baktoagar (Difco) 18,0 g Destilovaná voda 1,00 l
Složky se rozpustí a provede se sterilizace 0,5 l média v autoklávu při 115
°C po dobu 20 min.
b) Validovaná selektivní růstová média
Médium MTNA
Pokud není uvedeno jinak, pocházejí všechny složky médií z BDH.
Kvasnicový extrakt 2,0 g (Difco) Manit 2,5 g K2HPO40,25 g KH2PO40,25 g MgSO4. 7H2O 0,1 g MnSO4. H2O 0,015 g NaCl 0,05 g FeSO4. 7H2O 0,005 g Agar (Oxoid č. 1) 16,0 g Destilovaná voda 1,00 l
Složky se rozpustí, pH se upraví na 7,2. Po autoklávování (při 121 °C po
dobu 15 min.) a ochlazení na 50 st. C se přidají antibiotika: trimethoprim 0,06 g,
nalidixic acid 0,002 g, amphotericin B 0,01 g.
V zásobě se nechají roztoky antibiotik: trimethoprim (Sigma) a nalidixic
acid (Sigma) (obě 5 mg/ml) v 96 % metanolu, amphotericin B (Sigma) (1 mg/ml) v dimethyl
sulfoxidu. Zásobní roztoky jsou sterilizované filtrací.
Poznámka:
Trvanlivost základního média je 3 měsíce. Po přidání antibiotik
je trvanlivost 1 měsíc při skladování v chladu do 8 +/- 2 st. C.
Médium NCP-88 Živný agar (Difco) 23,0 g Kvasnicový extrakt 2,0 g (Difco) D-manit 5,0 g K2HPO42,0 g KH2PO40,5 g MgSO4. 7H2O 0,25 g Destilovaná voda 1,00 l
Složky se rozpustí,upraví se pH na 7,2. Po autoklávování a ochlazení na
50 °C se přidají následující antibiotika: polymyxin B sulphate (Sigma) 0,003g, nalidixic
acid (Sigma) 0,008 g, cycloheximide (Sigma) 0,2 g.
Antibiotika se rozpustí pro přípravu zásobních roztoků následovně: nalidixic
acid v 0,01 M NaOH, cykloheximid v 50% etanolu, polymyxin B sulphate v destilované
vodě. Zásobní roztoky se sterilizují filtrací.
Poznámka:
Trvanlivost základního média je 3 měsíce. Po přidání antibiotik
je trvanlivost 1 měsíc při skladování v chladu do 8 +/- 2 st. C.
Dodatek 6
Validované protokoly a činidla pro PCR
Poznámka:
Úvodní testování by mělo umožnit reprodukovatelné zjištění nejméně
10
3
až 104
buněk C. m. subsp. sepedonicus na 1 ml vzorkového extraktu. Úvodní testování
by také nemělo vykazovat žádné falešné pozitivní výsledky se skupinou vybraných kmenů
bakterií.1. Vícenásobný PCR protokol s interní PCR kontrolou (Pastrik, 2000)
1.1. Oligonukleotidní primery
primer PSA-1 5'- ctc ctt gtg ggg tgg gaa aa -3' primer PSA-R 5'- tac tga gat gtt tca ctt ccc c -3' primer NS-7-F 5'- gag gca ata aca ggt ctg tga tgc -3' primer NS-8-R 5’- tcc gca ggt tca cct acg ga -3’
Předpokládaná velikost amplikonu z DNA C. m. subsp. sepedonicus šablony =
502 bp (sada PSA-primerů).
Předpokládaná velikost amplikonu z interní PCR kontroly
18S rRNA = 377 bp (sada NS- primerů).
1.2. Reakční směs PCR
------------------------------------- ------------------------- ------------------------
Činidlo Množství na reakci Konečná koncentrace
------------------------------------- ------------------------- ------------------------
Sterilní voda (UPW) 15,725 µl
10x PCR pufr (1) (15 mM MgCl2) 2,5 µl 1x (1,5 mM MgCl2)
BSA (frakce V) (10 %) 0,25 µl 0,1 %
Směs d-nTP (20 mM) 0,125 µl 0,1 mM
Primer PSA-1 (10 µM) 0,5 µl 0,2 µM
Primer PSA-R (10 µM) 0,5 µl 0,2 µM
Primer NS-7-F (10 µM) (2) 0,1 µl 0,04 µM
Primer NS-8-R (10 µM) (2) 0,1 µl 0,04 µM
Taq polymeráza (5 U/µl) (1) 0,2 µl 1,0 U
Vzorkové množství 5,0 µl
------------------------------------- ------------------------- ------------------------
Celkový objem 25,0 µl
------------------------------------- ------------------------- ------------------------
(1) Metody byly validovány za použití Taq polymerázy Perkin Elmer (AmpliTaq
nebo Gold) a Gibco BRL.
(2) Koncentrace primerů NS-7 F a NS-8-R byla optimalizována pro extrakci
pupkové části bramboru za použití homogenizační metody a purifikace DNA podle Pastrika
(2000) (viz oddíl 6.1 písm. a) a 6.2). Při použití extrakce třepáním nebo jinými
metodami izolace DNA je nutná nová optimalizace koncentrací činidla.
1.3. Reakční podmínky pro PCR
Postupuje se podle následujícího programu:
1 cyklus: i) 3 minuty při teplotě 95 °C (denaturace DNA matrice)
10 cyklů ii) 1 minuta při teplotě 95 °C (denaturace DNA matrice)
iii) 1 minuta při teplotě 64 °C (připojení primerů)
iv) 1 minuta při teplotě 72 °C (prodlužování kopie)
25 cyklů v) 30 sekund při teplotě 95 °C (denaturace DNA matrice)
vi) 30 sekund při teplotě 62 °C (připojení primerů)
vii) 1 minuta při teplotě 72 °C (prodlužování kopie)
1 cyklus viii) 5 minut při teplotě 72 °C (závěrečná prodlužování)
ix) Udržuje se při teplotě 4 °C
Poznámka:
Tento program je optimalizován pro použití s tepelným cyklerem
MJ Research PTC 200. Při použití jiných modelů může být nutná modifikace kroků cyklů
ii), iii) iv), v), vi) a vii).
1.4. Analýzy ampliconu restriktivním enzymem
Produkty PCR amplifikované
z DNA C. m. subsp. sepedonicus produkují charakteristickou mnohotvárnost délky fragmentu
s enzymem Bgl II po inkubaci při teplotě 37 °C po dobu 30 minut. Fragmenty získané
z fragmentu specifického pro C. m. subsp. sepedonicus mají rozměry 282 bp a 220 bp.
2. Příprava nanášecího pufru
2.1. Bromfenolová modř (10 % zásobní roztok)
Bromfenolová modř 5 g
Destilovaná voda 50 ml
Nanášecí pufr
Glycerol (86 %) 3,5 ml
Bromfenolová modř 300 µl
Destilovaná voda 6,2 ml
3. Pufr 10x TRIS-acetát-EDTA (TAE), pH 8,0
TRIS 48,4 g Ledová kyselina octová 11,42 ml EDTA (sodná sůl) 3,72 g Destilovaná voda 1,00 l
Před použitím se zředí na 1x.
Také komerčně dostupné (např. Invitrogen nebo rovnocenné).
Dodatek 7
Validovaná činidla pro FISH test
1. Oligosondy
Sonda specifická pro Cms CMS-CY3-01: 5’- ttg cgg ggc gca cat ctc tgc acg -3’ Nespecifická eubakteriální sonda EUB-338-FITC: 5’- gct gcc tcc cgt agg agt-3’
2. Fixační roztok
[UPOZORNĚNÍ: FIXAČNÍ ROZTOK OBSAHUJE PARAFORMALDEHYD, KTERÝ JE TOXICKÝ!
POUŽÍVAT RUKAVICE A NEVDECHOVAT. DOPORUČUJE SE PRACOVAT V DIGESTOŘI.]
i) Zahřeje se 9 ml molekulárně čisté vody (např. Ultra pure water = (UPW))
na teplotu přibližně 60 °C a přidá se 0,4 g paraformaldehydu. Paraformaldehyd se
rozpustí po přidání 5 kapek 1N NaOH a zamíchání magnetickým míchadlem.
ii) Upraví se pH na 7,0 přidáním 1ml fosfátového pufru 0,1 M (PB; pH 7,0)
a 5 kapek HCl 1N. Indikačním proužkem se zkontroluje pH a v případě potřeby se upraví
pomocí HCl nebo NaOH.
[UPOZORNĚNÍ: V ROZTOCÍCH S PARAFORMALEDHYDEM NEPOUŽÍVAT MĚŘIČ pH!]
iii) Roztok se přefiltrujte přes membránový filtr 0,22 µm a skladuje se chráněný
před prachem při teplotě 4 °C do dalšího použití.
iv) Poznámka:
Alternativní fixační roztok: 96 % etanol
.3. 3x Hybmix
NaCl 2,7 M Tris-HCl 60 mM (pH 7,4) EDTA (sterilizovaný přes filtr a 15 mM autoklávovaný)
Zředí se až 1x, podle potřeby.
4. Hybridizační roztok
1x Hybmix Sodium dodecyl sulfát (SDS) 0,01 % Sonda EUB 338 5 ng/mikrol Sonda CMSCY301 5 ng/mikrol
Připraví se množství hybridizačního roztoku podle výpočtů v tabulce. Pro
každé sklíčko (obsahující dvojmo 2 různé vzorky) je třeba 90 mikrol hybridizačního
roztoku.
Tabulka: Doporučená množství pro přípravu hybridizační směsi
------------------------------------- -------------------------
2 sklíčka 8 sklíček
------------------------------------- -------------------------
Sterilní ultra čistá voda 50,1 200,4
3x hybmix 30,0 120,0
1 % SDS 0,9 3,6
sonda EUB 338 (100 ng/mikrol) 4,5 18,0
sonda CMSCY301 (100 ng/mikrol) 4,5 18,0
------------------------------------- -------------------------
Celkový objem (mikrol) 90,0 360,0
------------------------------------- -------------------------
Poznámka.:
Všechny roztoky obsahující světlocitlivé oligosondy se uchovávají
v temnu při teplotě - 20 °C. Během použití je nutné je ochraňovat před přímým slunečním
zářením nebo elektrickým světlem.
5. 0,1M fosfátový pufr, pH 7,0
Na2HPO48,52 g KH2PO45,44 g Destilovaná voda 1,001
Složky se rozpustí, zkontroluje se pH a sterilizuje se autoklávováním při
teplotě 121 °C po dobu 15 minut.
Dodatek 8
Pěstování lilku vejcoplodého
Semena lilku vejcoplodého (Solanum melongena) se vysejí do pasterizovaného
výsevního substrátu. Semenáčky s plně rozvinutými děložními lístky (10 až 14 dní)
se přepichují do pasterizovaného pěstebního substrátu.
Lilek by se měl pěstovat ve skleníku za následujících podmínek:
Délka dne: 14 hodin nebo přirozená délka dne, pokud je delší;
Teplota: den: 21 až 24 °C,
noc: 15 °C.
Vhodné odrůdy lilku vejcoplodého
„Black Beauty“,
„Long Tom“,
„Rima“,
„Balsas“.
Dodavatel: viz internetová stránka
http://forum.europa.eu.int/Public/irc/sanco/Home/main
Dodatek 9
Gramovo barvení v Huckerově modifikaci (Doetsch, 1981)(1)
Roztok krystalové violeti
Rozpustí se 2 g krystalové violeti v 20 ml 95 % etanolu.
Rozpustí se 0,8 g šťavelanu amonného v 80 ml destilované vody. Oba roztoky
se smíchají.
Lugolův jódový roztok Jód 1 g Jodid draselný 2 g Destilovaná voda 300 ml
Pevné látky se společně rozetřou pomocí tloučku v misce. Nasypou se do vody
a míchají se v uzavřené nádobě do rozpuštění.
Safraninový roztok kontrastního barviva
Zásobní roztok: Safranin O 2,5 g 95 % etanol 100 ml
Zamíchá se a uloží se.
Ředění: 1:10 pro přípravu pracovního roztoku.
Postup barvení
1. Připraví se roztěry, vysuší se na vzduchu a fixují se zahřátím.
2.
Sklíčko se zalije roztokem krystalové violeti a nechá se působit 1 minutu.
3. Krátce
se omyje pod tekoucí vodou.
4. Zalije se Lugolovým jódovým roztokem a nechá se působit
po dobu jedné minuty.
5. Opláchne se pod tekoucí vodou a vysuší se savým papírem.
6.
Odbarvuje se pomocí po kapkách přidávaného 95 % etanolu tak dlouho, pokud se vyplavuje
barvivo, nebo ponořením za jemného pohybování do etanolu na dobu 30 sekund.
7. Opláchne
se pod tekoucí vodou a vysuší se savým papírem.
8. Zalije se safraninovým roztokem
a nechá se působit 10 s.
9. Opláchne se pod tekoucí vodou a vysuší se savým papírem.
Grampozitivní bakterie se zbarví fialově modře, gramnegativní bakterie se
zbarví růžovočerveně.
(1) Mohou se také použít komerčně dostupné roztoky nebo barvicí soupravy.
LITERATURA
1. Anonymous, 1987. Scheme of the detection and diagnosis of the ring rot
bacterium Corynebacterium sepedonicum in batches of potato tubers. Commission of
the European Communities, Luxembourg. Publ EUR 11 288 EN, 21 pp.
2. Bradbury, J. F., 1970. Isolation and preliminary study of bacteria from
plants. Rev. Pl. Path., 49, 213-218.
3. Dinesen, I. G., 1984. The extraction and diagnosis of Corynebacterium
sepedonicum from diseased potato tubers. EPPO Bull. 14 (2), 147-152.
4. Doetsch, R. N., 1981. Determinative methods of light microscopy. V:
Manual of methods for general bacteriology, American Society for Microbiology, Washington,
21-23.
5. Hugh, R. a Leifson, F., 1953. The taxonomic significance of fermentative
versus oxidative metabolism of carbohydrates by various gram-negative bacteria. J.
Bact., 66, 24-26.
6. Janse, J. D., 1991. Infra- and intra-specific classification of Pseudomonas
solanacearum strains using whole cell fattyacid analysis. Systematic and Applied
Microbiology 14; 335-345.
7. Janse, J. D. a J. Van Vaerenbergh. The interpretation of the EC method
for the detection of latent ring rot infections (Corynebacterium sepedonicum) in
potato. EPPO Bull., č. 17, 1987, pp. 1-10.
8. Jansing, H. a K. Rudolph, 1998. Physiological capabilities of Clavibacter
michiganensis subsp. sepedonicus and development of a semi-selective medium. Journal
of Plant Diseases and Protection, 105, 590-601.
9. Kovacs, N., 1956. Identification of Pseudomonas pyocyanea by the oxidase
reaction. Nature, Lond., 178, 703.
10. Klement Z.; Rudolph, K a D. C. Sands, 1990. Methods in Phytobacteriology.
Akadémiai Kiadó, Budapest, 568 pp.
11. Lelliott, R. A., 1966. The plant pathogenic coryneform bacteria. J.
appl. Bact., 29, 114-118.
12. Lelliott, R. A., E. Billing a A. C. Hayward, 1966. A determinative
scheme for the fluorescent plant pathogenic pseudomonads J. appl. Bact., 29, 470-489.
13. Lelliott, R. A. a P. W., Sellar, 1976. The detection of latent ring
rot (Corynebacterium sepedonicum (Spiek. et Kotth.) Skapt. et Burkh.) in potato stocks.
EPPO Bull., 6 (2), 101-106.
14. Li, X. a S.H. de Boer, 1995. Selection of Polymerase Chain Reaction
primers from RNA intergenic spacer region for specific detection of Clavibacter michiganensis
subsp. sepedonicus. Phytopathology, 85, 837-842.
15. Mills, D., Russell, B., W. a J., W. Hanus, 1997. Specific detection
of Clavibacter michiganensis subsp. sepedonicus by amplification of three unique
DNA sequences isolated by subtraction hybridization. Phytopathology, 87, 8, 853-861.
16. Pastrik, K. -H. a R.A. Rainey. 1999. Identification and differentiation
of Clavibacter michiganensis subspecies by polymerase chain reaction-based techniques.
J. Phytopathology 147; 687-693.
17. Pastrik, K.-H., 2000. Detection of Clavibacter michiganensis subsp.
sepedonicus in potato tubers by multiplex PCR with coamplification of host DNA. European
Journal of Plant Pathology, 106, 155-165.
18. Ramamurthi, C. S., 1959. Comparative studies on some Gram-positive
phytopathogenic bacteria and their relationship to the Corynebacteria. Mem. Cornell
agric. Exp. Sta., 366, 52 pp.
19. Schaad, W., Berthier-Schaad, Y., Sechler, A. a Knorr, D. (1999) Detection
of Clavibacter michiganensis subsp. sepedonicus in potato tubers by BIO-PCR and an
automated real-time fluorescence detection system. Plant Disease 83; 1095-1100.
20. Schaad, W. 2001. Laboratory guide for identification of plant pathogenic
bacteria. Schaad [Hrsg.]. - 3. ed.; St. Paul, Minnesota:, 373 pp.
21. Skerman, V. B. D., 1967. A guide to the identification of the genera
of bacteria. 2nd ed., William and Wilkins Company, Baltimore.
22. Smith, N. C.; Hennesy, J; Stead, D.E., 2001. Repetetive sequence-derived
PCR profiling using the BOX-A1 Ralstonia solanacearum primer for rapid identification
of plant pathogen Clavibacter michiganensis subsp. sepedonicus. European Journal
of Plant Pathology, 107 (7), 739-748.
23. Sneath, P. H. A. and V. G. Collins, 1974. A study in test reproductibility
between laboratories: report of Pseudomonas working party. Antonie van Leeuwenhoek,
40, 481-527.
24. Stead, D.E. 1992. Grouping of plant pathogenic and some other Pseudomonas
spp. using cellular fatty-acid profiles. International Journal of Systematic Bacteriology
42; 281-295.
25. Wullings, B. A.; van Beuningen, A. R.; Janse, J. D. a A. D. L. Akkermans,
1998. Detection of Ralstonia solanacearum, which causes brown rot of potato, by fluorescent
in situ hybridization with 23s rRNA-targeted probes. Appl. Environ. Microbiol. 64,
4546-4554.
B. METODY DIAGNÓZY, DETEKCE A IDENTIFIKACE PŮVODCE HNĚDÉ HNILOBY
Předložené postupové diagramy popisují různé postupy, které jsou součástí:
i) diagnózy hnědé hniloby v hlízách bramboru a bakteriálního vadnutí bramboru,
rajčete a některých dalších hostitelských rostlin,
ii) detekce Ralstonia solanacearum ve vzorcích hlíz bramboru, rostlin
bramboru, rostlin rajčete a jiných hostitelských rostlin, a ve vzorcích vody a půdy,
iii) identifikace Ralstonia solanacearum (R. solanacearum)
OBECNÉ ZÁSADY
Optimalizované protokoly pro různé metody, schválená činidla a podrobnosti
pro přípravu testovaných a kontrolních materiálů jsou uvedeny v dodatcích. Seznam
laboratoří, které se podílely na optimalizaci a validaci protokolů, je v dodatku
1.
Protože protokoly obsahují zjištění karanténního organismu a zahrnují použití
životaschopných kultur R. solanacearum jako kontrolních materiálů, je nutné pracovat
za vhodných karanténních podmínek s odpovídajícím zařízením na odstraňování odpadů
a za podmínek stanovených příslušným povolením Ústavu.
Testovací parametry musí zajišťovat stálé a reprodukovatelné zjištění úrovní
R. solanacearum jako stanovené prahy vybraných metod.
Zcela nezbytná je přesná příprava pozitivních kontrol.
Testování podle požadovaných prahů také zahrnuje správné nastavení, údržbu
a kalibraci zařízení, pečlivé zacházení s činidly a jejich uchovávání a všechna opatření
pro zamezení kontaminace mezi vzorky, např. oddělení pozitivních kontrol od testovaných
vzorků. Musí být uplatněno standardní řízení kvality, aby se zabránilo administrativním
a jiným chybám, zvláště při označování a v dokumentaci.
Podezření z výskytu, jak je uvedeno v § 4 odst. 1, naznačuje pozitivní výsledek
z diagnostických nebo screeningových testů provedených na vzorku, jak je znázorněno
v níže uvedených postupových diagramech. První pozitivní screeningový test ( test
IF, PCR/FISH, selektivní izolace) musí být potvrzen druhým screeningovým testem založeném
na jiném biologickém principu.
Pokud je první screeningový test pozitivní, existuje podezření na infekci
R. solanacearum a musí být proveden druhý screeningový test. Pokud je i druhý screeningový
test pozitivní, pak je podezření potvrzeno a musí se pokračovat v testování podle
daného schématu. Jestliže je druhý screeningový test negativní, pak vzorek není považován
za infikovaný R. solanacearum.
Potvrzená přítomnost podle §. 5 odst. 1 vyžaduje izolaci a identifikaci čisté
kultury R. solanacearum s potvrzením patogenity.
1. Použití postupových diagramů
1.1. Postupový diagram pro diagnózu hnědé hniloby a bakteriálního vadnutí
(Ralstonia solanacearum) hlíz bramboru a rostlin bramboru, rajčete a jiných hostitelských
rostlin vykazujících příznaky hnědé hniloby nebo bakteriálního vadnutí
Postup testování je určen pro hlízy a rostliny bramboru s podezřením z výskytu
nebo typickými příznaky hnědé hniloby nebo bakteriálního vadnutí. Zahrnuje rychlý
screeningový test, izolaci patogena z infikovaného vodivého pletiva na selektivní
živné půdě a v případě pozitivního výsledku identifikaci kultury Ralstonia solanacearum.

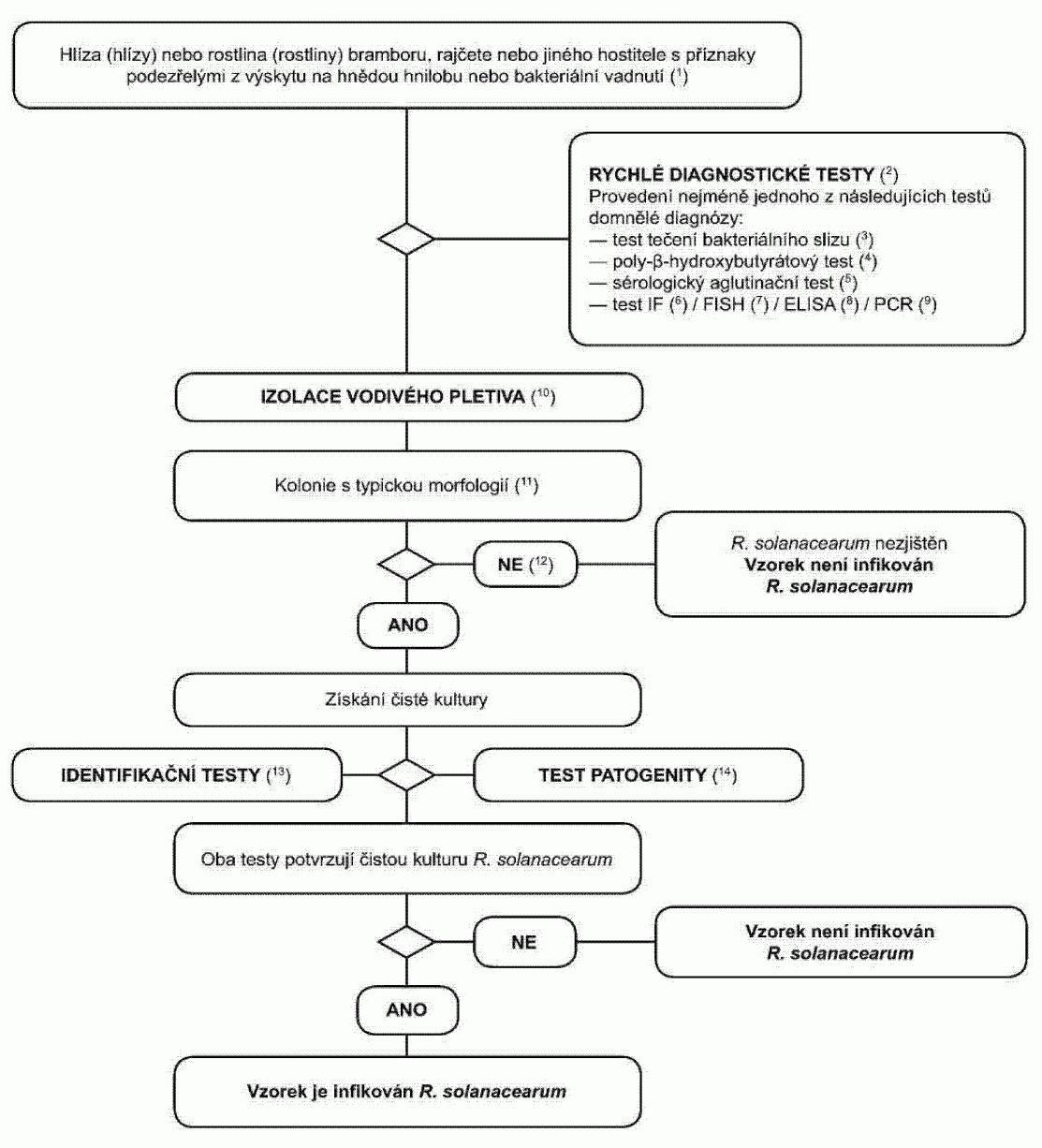
-----
1) Popis příznaků se nachází v oddíle 2.1.
2) Rychlé diagnostické testy usnadňují předběžnou diagnózu, ale nejsou
dokonalé. Negativní výsledek naznačuje vždy nepřítomnost patogenu.
3) Test na výtok slizu ze svazků cévních stonku je popsán v oddíle 6.1.1.
4) Test na přítomnost granulí poly-beta-hydroxybutyrátu je popsán v oddíle
6.1.2.
5) Sérologické aglutinační testy bakteriálního slizu nebo extraktů z
pletiva vykazujícího příznaky jsou popsány v oddíle 6.1.3.
6) Test IF bakteriálního slizu suspendovaného ve vodě nebo extraktech
pletiva vykazujícího příznaky je popsán v oddíle 6.1.5.
7) Test FISH bakteriálního slizu suspendovaného ve vodě nebo extraktech
pletiva vykazujícího příznaky je popsán v oddíle 6.1.7.
8) Test ELISA bakteriálního slizu suspendovaného ve vodě nebo extraktech
pletiva vykazujícího příznaky je popsán v oddíle 6.1.8.
9) Test PCR bakteriálního slizu suspendovaného ve vodě nebo extraktech
pletiva vykazujícího příznaky je popsán v oddíle 6.1.6.
10) Patogen je obvykle snadno izolovatelný z rostlinného materiálu vykazujícího
příznaky metodou zřeďovacích roztěrů (postupným ředěním) (viz. v oddíle 2.3.).
11) Typická morfologie kolonie je popsána v oddíle 2.3.d.
12) Kultivace může selhat při pokročilých fázích infekce z důvodů konkurence
nebo bujného růstu saprofytických bakterií. Jestliže jsou příznaky infekce typické,
ale izolační test je negativní, musí se izolace opakovat, nejlépe kultivací na selektivních
živných půdách.
13) Spolehlivá identifikace čistých kultur R. solanacearum se provede
pomocí testů popsaných v bodu 6.2. Dílčí specifická charakteristika je nepovinná,
ale doporučuje se pro každý nový případ.
14) Test patogenity je popsán v bodu 6.3.
1.2. Postupový diagram pro detekci a identifikaci Ralstonia solanacearum
ve vzorcích hlíz bramboru nevykazujících příznaky
Testování je určeno ke zjištění latentních infekcí v hlízách bramboru. Pozitivní
výsledek alespoň ze dvou screeningových testů (viz poznámka v diagramu 3), z nichž
každý je založen na jiném biologickém principu, musí být doplněn izolací patogenu
a dále, v případě izolace typických kolonií, potvrzením, že čistá kultura je R. solanacearum.
Pozitivní výsledek pouze z jednoho screeningového testu není dostačující k tomu,
aby byl vzorek považován za podezřelý. Screeningové testy a izolační testy musí umožnit
detekční práh 10
3
až 104
buněk/ml resuspendované pelety zahrnutý jako pozitivní kontroly
v každé sérii testů.

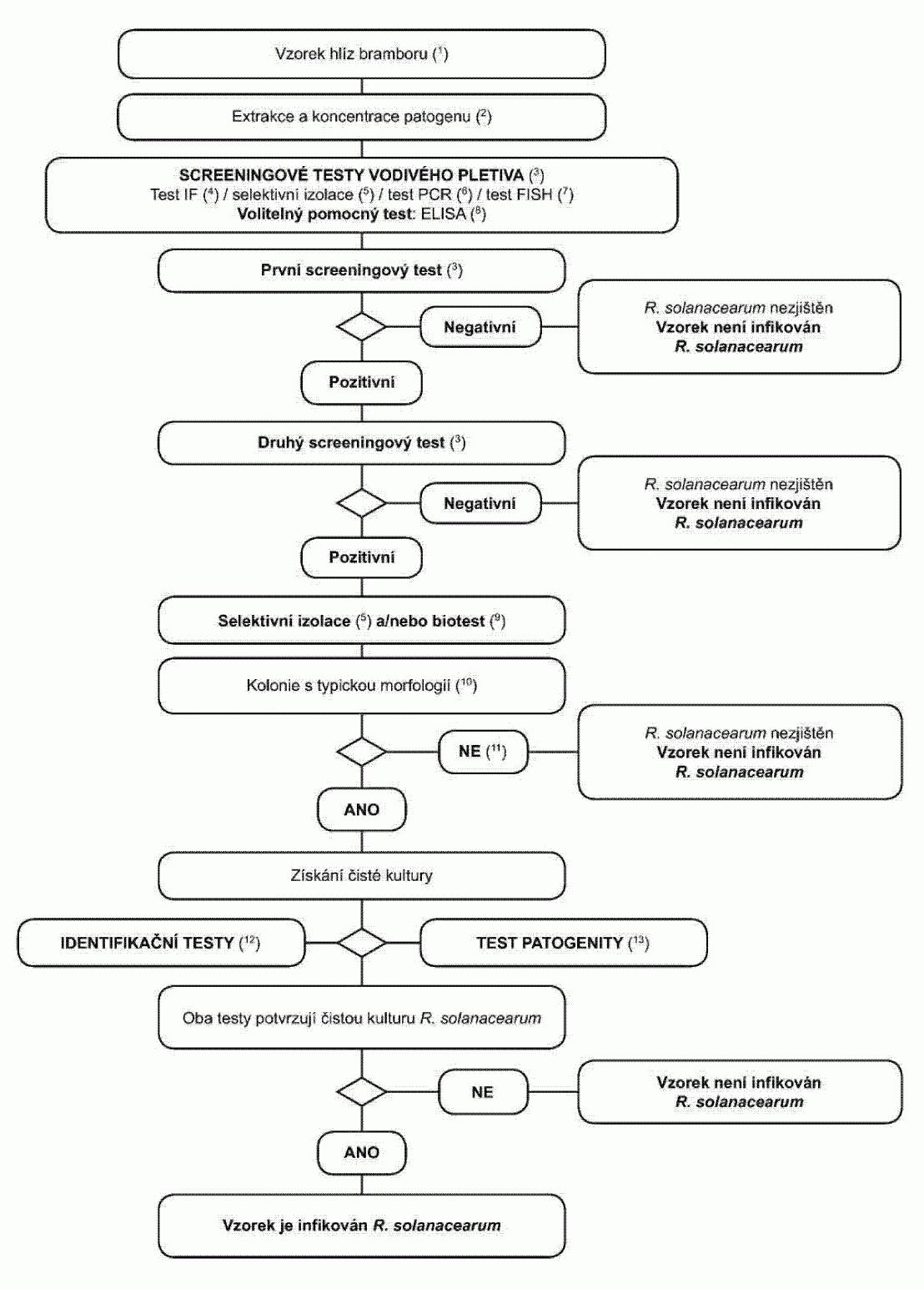
-----
1) Standardní velikost vzorku je 200 hlíz, ačkoli postup lze použit i
na menší počet, jestliže 200 hlíz není k dispozici.
2) Metody extrakce a koncentrace patogenu jsou popsány v oddíle 3.1.1.
3) Pokud jsou alespoň dva testy založené na různých biologických principech
pozitivní, musí být provedena izolace a potvrzení. Provede se nejméně jeden screeningový
test. Je-li test negativní, je vzorek považován za negativní. V případě, že je test
pozitivní, je pro validaci prvního pozitivního výsledku nezbytný ještě jeden nebo
více screeningových testů založených na různých biologických principech. Je-li druhý
nebo další test negativní, je vzorek považován za negativní. Další testy nejsou nutné.
4) Test IF je popsán v oddíle 6.1.5.
5) Test selektivní izolace je popsán v oddíle 6.1.4.
6) Testy PCR jsou popsány v oddíle 6.1.6.
7) Test FISH je popsán v oddíle 6.1.7.
8) Testy ELISA jsou posány v oddíle 6.1.8.
9) Biotest je popsán v oddíle 6.1.9.
10) Typická morfologie kolonie je popsána v oddíle 2.3.d.
11) Kultivace nebo biotest mohou selhat z důvodů konkurence nebo inhibice
saprofytickými bakteriemi. Jsou-li ve screeningových testech získány pozitivní výsledky,
ale izolační testy jsou negativní, je třeba opakovat izolační testy ze stejné pelety
nebo dodatečným odebráním vodivého pletiva z blízkosti pupkového konce hlíz stejného
vzorku, případně provést test s dalšími vzorky.
12) Spolehlivé identifikace čistých kultur podezřelých na R. solanacearum
se dosáhne pomocí testů popsaných v oddíle 6.2.
13) Test patogenity je popsán v oddíle 6.3.
Postupový diagram pro detekci a identifikaci Ralstonia solanacearum ve vzorcích
rostlin bramboru nevykazujících příznaky, rostlin rajčete nevykazujících příznaky
nebo rostlin jiných hostitelských rostlin nevykazujících příznaky

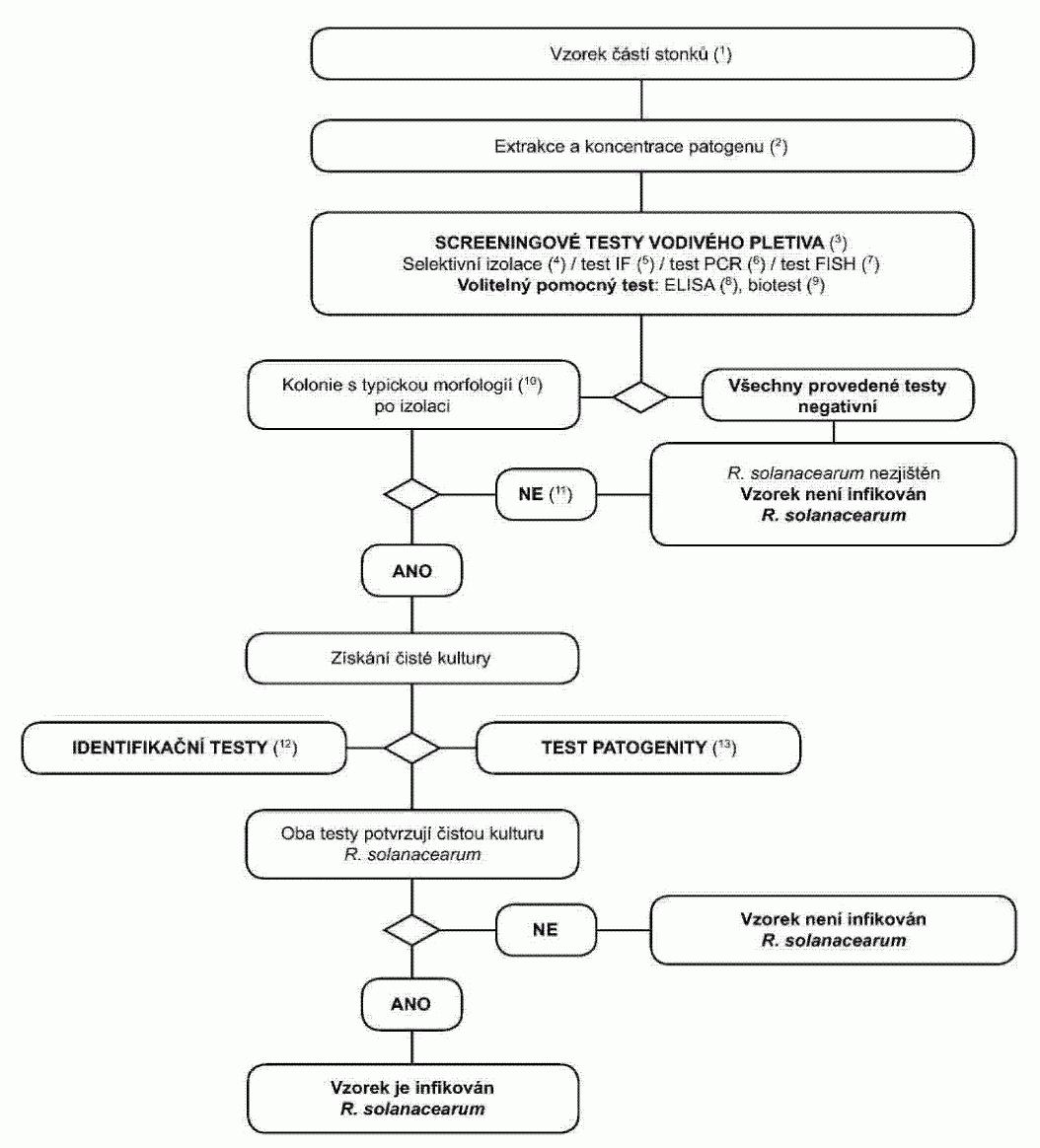
-----
1) Viz. oddíl 3.2.1., kde jsou uvedeny doporučené velikosti vzorků.
2) Metody extrakce a koncentrace patogenu jsou popsány v oddíle 3.2.1.
3) Pokud jsou alespoň dva testy založené na různých biologických principech
pozitivní, musí být provedena izolace a potvrzení. Provede se alespoň jeden screeningový
test. Je-li test negativní, je vzorek považován za negativní. V případě, že je test
pozitivní, vyžaduje se pro validaci prvního pozitivního výsledku provedení druhého
nebo více screeningových testů založených na různých biologických principech. Jsou-li
druhý nebo další testy negativní, považuje se vzorek za negativní. Další testy nejsou
nutné.
4) Selektivní izolační test je popsán v oddíle 6.1.4.
5) Test IF je popsán v oddíle 6.1.5.
6) Testy PCR jsou popsány v oddíle 6.1.6.
7) Test FISH je popsán v oddíle 6.1.7.
8) Testy ELISA jsou popsány v oddíle 6.1.8.
9) Biotest je popsán v oddíle 6.1.9.
10) Typická morfologie kolonie je popsána v oddíle 2.3.d.
11) Kultivace nebo biotest mohou selhat z důvodů konkurence nebo inhibice
saprofytickými bakteriemi. Jsou-li ve screeningových testech získány pozitivní výsledky,
ale izolační testy jsou negativní, opakují se izolační testy.
12) Spolehlivé identifikace čistých kultur, u kterých je podezření, že
se jedná o R. solanacearum, se dosáhne pomocí testů popsaných v oddíle 6.2.
13) Test patogenity je popsán v oddíle 6.3.
2. Diagnóza původce hnědé hniloby
2.1. Příznaky
2.1.1. Příznaky u bramboru
Rostlina bramboru.
Raná fáze infekce v polních
podmínkách se rozpozná vadnutím listů směrem k vrcholu rostliny při vysokých teplotách
během dne, přičemž v noci dochází k zotavení. V raných fázích vadnutí zůstávají listy
zelené, ale později žloutnou a objevují se hnědé nekrózy. Dochází také k ohýbání
listů dolů. Vadnutí jednoho výhonku nebo celé rostliny se rychle stává nevratným
a končí kolapsem a uhynutím rostliny. Z cévních svazků napříč uříznutých stonků zvadlých
rostlin obvykle vytéká hnědý a mléčný bakteriální sliz nebo je možné sliz vymáčknout.
Při ponoření uříznutého stonku svisle do vody se z cévních svazků táhnou vlákna slizu.
Hlíza bramboru. Hlízy bramboru je třeba překrojit napříč u pupkového konce a podélně
přes pupkový konec. V rané fázi se infekce pozná podle láhvově žlutého až světle
hnědého zabarvení cévního prstence, ze kterého po několika minutách začne prýštit
bledý krémový bakteriální sliz. Později se cévní zbarvení stává výrazněji hnědým
a odumření se může rozšířit do parenchymatického pletiva. V pokročilejších fázích
se infekce rozšíří vně od pupkového konce hlízy a z oček může vytékat bakteriální
sliz, na který se přilepují částice půdy. Na slupce se mohou objevit červenohnědá,
lehce propadlá místa jako důsledek vnitřního kolapsu cévního pletiva. V pokročilejších
fázích infekce je obvyklý sekundární rozvoj měkkých hnilob bakteriálního a houbového
původu.
2.1.2. Příznaky u rajčete
Rostlina rajčete. Prvním viditelným příznakem
je povadlý vzhled nejmladších listů. Za příznivých podmínek pro patogena (teplota
půdy kolem 25 st. C, při nasycené vzdušné vlhkosti) se během několika málo dní rozvine
kroucení listů směrem dolů a vadnutí na jedné straně rostliny nebo celé rostliny,
které končí jejím úplným odumřením. Za méně příznivých podmínek pro patogena (teplota
půdy méně než 21şC) rostlina tolik nevadne, ale na stonku se může tvořit větší počet
postranních výhonů. Je možné pozorovat vodou nasáklé pruhy od spodu stonku, které
jsou dokladem odumírání cévního systému. Při příčném řezu stonkem vylučují hnědě
zbarvená vodivá pletiva bílý nebo nažloutlý bakteriální sliz.
2.1.3. Příznaky u jiných hostitelů
Rostliny Solanum dulcamara a S. nigrum.
Za normálních podmínek jsou u těchto plevelných hostitelských rostlin zřídkakdy pozorovány
příznaky vadnutí, pokud teploty půdy nepřevyšují 25 st. C nebo není extrémně vysoká
koncentrace inokula (např. u rostliny S. nigrum rostoucí u nemocné rostliny bramboru
nebo rajčete). Při vadnutí jsou příznaky stejné jako u rostliny rajčete. Nevadnoucí
rostliny S. dulcamara, která má stonky a kořeny ve vodě, mohou vykazovat vnitřní
světle hnědé zbarvení vodivých pletiv na příčném řezu spodní části stonku nebo částí
stonku pod vodou. Z řezu cévních svazků mohou vytékat bakterie nebo mohou tvořit
vlákna slizu, jestliže je řez stonku ponořen svisle do vody, a to i při absenci příznaků
vadnutí.
2.2. Rychlé screeningové testy
Rychlé screeningové testy mohou napomoci předběžné
diagnóze, ale nejsou dostačující. Použije se jeden nebo více následujících testů:
2.2.1. Test na výtok slizu ze stonku (Viz. 6.1.1.)
2.2.2. Test na přítomnost granulí poly-beta- hydroxybutyrátu (PHB)
Charakteristické
granule PHB v buňkách R. solanacearum jsou zviditelněny obarvením tepelně fixovaných
skvrn bakteriálního slizu z infikovaného pletiva na mikroskopickém sklíčku nilskou
modří A nebo súdánskou černí (viz. oddíl 6.1.2.).
2.2.3. Sérologické aglutinační testy
(Viz. oddíl 6.1.3.)
2.2.4. Jiné testy
Dalšími vhodnými rychlými screeningovými testy jsou test
IF (viz.oddíl 6.1.5.), test FISH (viz.oddíl 6.1.7.), testy ELISA (viz.oddíl 6.1.8.)
a testy PCR (viz. oddíl 6.1.6.).
2.3. Postup při izolaci
a) Odebere se sliz nebo vrstva zbarveného pletiva z cévního prstence hlízy
bramboru nebo z cévních vláken stonku rostliny bramboru, rajčete nebo jiné vadnoucí
hostitelské rostliny. Suspenduje se v malém množství sterilní destilované vody nebo
50mM fosfátového pufru (dodatek 4) a nechá se 5-10 minut stát.
b) Připraví se řada desetinásobných ředění suspenze.
c) Přenese se 50-100 ěl suspenze a roztoku na universální živnou půdu
(NA, YPGA neboSPA; viz. dodatek 2) a/nebo Kelmanovo tetrazolové médium (dodatek 2)
a/nebo validované selektivní médium (např. SMSA, viz. dodatek 2). Rozetře se metodou
zřeďovacích roztěrů. Připraví se případně samostatné misky s rozředěnou buněčnou
suspenzí R. solanacearum biovar 2 k pozitivní kontrole.
d) Inkubuje se 2-6 dní při teplotě 28 st. C.
- Na universální živné půdě
vytvářejí virulentní izoláty R. solanacearum perlově krémově bílé, ploché, nepravidelné
a fluidní kolonie, často s charakteristickými spirálkami ve středu. Aviruletní formy
R. solanacearum tvoří malé kruhové nefluidní máslovité kolonie, které jsou zcela
krémově bílé.
- U Kelmanova tetrazolového média a média SMSA jsou spirálky krvavě
červeně zbarvené. Aviruletní formy R. solanacearum tvoří malé kruhové nefluidní máslovité
kolonie, které jsou zcela temně červené.
2.4 Identifikační testy Ralstonia solanacearum
Testy potvrzující výskyt
R. solanacearum v podezřelých izolátech jsou popsány v bodu 6.2.
3. Detekce a identifikace původce hnědé hniloby ve vzorcích hlíz bramboru
3.1. Podrobné metody pro detekci a identifikaci Ralstonia solanacearum ve
vzorcích bezpříznakových hlíz bramboru
3.1.1. Příprava vzorků
Poznámka:
- Standardní velikost vzorku je 200 hlíz na jeden test. Intenzivnější
vzorkování vyžaduje více testů na vzorcích této velikosti. Větší množství hlíz ve
vzorku vede k zpomalení nebo složitějšímu výkladu výsledků. Postup lze vhodně použít
i pro vzorky s méně než 200 hlízami, pokud je k dipozici menší množství hlíz.
- Validace
všech níže uvedených zjišťovacích metod je založená na testování vzorků o velikosti
200 hlíz.
- Extrakt bramboru popsaný níže může být rovněž použit pro zjištění původce
bakteriální kroužkovitosti bramboru. Clavibacter michiganensis subsp. sepedonicus.
Nepovinné ošetření před přípravou vzorku
a) Inkubace vzorků při teplotě 2-30 st. C po dobu až 2 týdnů před testováním
na podporu množení všech populací R. solanacearum.
b) Hlízy se omyjí. Použijí se vhodné desinfekční prostředky (s obsahem
chlóru, jestliže má být proveden PCR test, pro odstranění patogenní DNA) a mycí prostředky
mezi každým vzorkem. Hlízy se nechají oschnout na vzduchu. Tento postup mytí je vhodný
zvláště v případě, když je ve vzorcích příliš zeminy nebo jestliže má být proveden
test PCR nebo přímá izolace.
3.1.1.1. Pomocí čistého a dezinfikovaného skalpelu nebo nože na zeleninu
se odstraní slupka na pupkovém konci každé hlízy, tak aby bylo viditelné vodivé pletivo.
Opatrně se vyříznou malé výkrojky vodivých pletiv na pupkovém konci. Množství pletiva
nezahrnující cévní svazky se omezí na minimum.
Poznámka:
Všechny (hnijící) hlízy s podezřelými příznaky hnědé hniloby
se dají stranou a testují se odděleně.
Pokud jsou při vyříznutí výkrojku zjištěny
příznaky podezření na hnědou hnilobu, provede se vizuální vyšetření takové hlízy
na řezu hlízy na pupkovém konci. Všechny naříznuté hlízy s podezřelými příznaky se
uchovávají po dobu nejméně 2 dnů při pokojové teplotě, aby mohlo dojít ke zkorkovatění
(suberizaci) a potom se uchovávají v chladu (4-10 st. C) za řádných karanténních
podmínek. Všechny hlízy včetně hlíz s podezřelými příznaky by měly být uchovány podle
přílohy III.
3.1.1.2. Výkrojky z pupkové části se uloží do nepoužitých nádob na jedno
použití, které jsou uzavíratelné a/nebo utěsnitelné (v případě, že nádoby jsou znovu
používány, musí být důkladně vyčištěny a dezinfikovány prostředky s obsahem chlóru).
Nejlépe je zpracovat výkrojky z pupkového konce okamžitě. Není-li to možné, uchovávají
se v nádobě bez přidání pufru; v chladu nejdéle 72 hodin nebo při pokojové teplotě
(18 - 25 st. C) nejdéle 24 hodin.
Výkrojky z pupkových konců se zpracují jedním z
následujících postupů:
a) Zalijí se dostatečným množství (přibližně 40 ml) extrakčního pufru (dodatek
4) a třepou se v rotační třepačce (50-100 ot./min) 4 hodiny při teplotě nižší než
24 st. C nebo 16-24 hodin chlazené, nebo
b) homogenizují se dostatečným množství (přibližně 40 ml) extrakčního pufru
(dodatek 4) buď v mixéru (např. Waring nebo Ultra Thurax) nebo drcením v utěsněném
maceračním sáčku na jedno použití (např. silný polyethylenový sáček Stomacher nebo
Bioreba, 150mm x 250mm, sterilovaný zářením) pomocí gumového tloučku nebo vhodného
mlecího zařízení (např. Homex).
Poznámka:
Při homogenizaci vzorků pomocí mixéru existuje vysoké nebezpečí
křížové kontaminace vzorků. Je nutné zamezit vzniku aerosolu nebo rozlití během extrakčního
procesu. Pro každý vzorek se použijí čerstvě sterilované nože (ostří) a nádoby. Při
testu PCR je nutno zabránit přenosu DNA na nádobách nebo v mlecím přístroji. U testu
PCR se doporučuje použít drcení v sáčcích na jedno použití a použití zkumavek na
jedno použití.
3.1.1.3. Supernatant se dekantuje. Je-li příliš zakalený, pročistí se buď
pomalým odstřeďováním (nejvýš 180 g po dobu 10 minut při teplotě 4-10 st. C) nebo
vakuovou filtrací (40-100µm), filtr se omyje přídavkem (10ml) extrakčního pufru.
3.1.1.4. Bakteriální frakce se zahustí odstřeďováním, 7 000g po dobu 15
minut (nebo 10 000 g po dobu 10 minut) při teplotě 4-10 st. C a odstraní se supernatant,
aniž by se rozvířila peleta.
3.1.1.5. Peleta se resuspenduje v 1,5 ml peletového pufru (dodatek 4). Použije
se 500 µl pro R. solanacearum, 500 µl pro Clavibacter michiganensis subsp. sepedonicus
a 500 µl pro referenční účely. Přidá se sterilní glycerol na konečnou koncentraci
10 - 25% (v/v) k 500 µl referenční poměrné části a ke zbývající části vzorku, promíchá
se vířením a uloží se při teplotě -16 až -24 st. C (týdny) nebo - 68 až -86 st. C
(měsíce). Extrakty se uchovávají během testování při teplotě 4-10 st. C.
Opakované zmrazení a rozmrazení se nedoporučuje.
Je-li nutná přeprava extraktu, zajistí se přeprava v chladícím boxu s doručením
do 24 až 48 hodin.
3.1.1.6. Je nezbytně nutné, aby všechny pozitivní kontroly a vzorky R. solanacearum
byly uchovány a zpracovány odděleně, aby se zabránilo vzájemné kontaminaci. To platí
pro sklíčka IF a všechny testy.
3.1.2. Testování
Postupové diagramy a popisy testů a optimalizovaných protokolů jsou v příslušných
dodatcích:
Selektivní izolace (viz. oddíl 6.1.4.)
IF test (viz.oddíl 6.1.5.)
Testy PCR
(viz. oddíl 6.1.6.)
Test FISH (viz. oddíl 6.1.7.)
Testy ELISA (viz. oddíl 6.1.8.)
Biotest
(viz. oddíl 6.1.9.)
3.2. Podrobné metody pro detekci a identifikaci R. solanacearum ve vzorcích
bezpříznakových rostlin bramboru, rajčete nebo jiných hostitelských rostlin
3.2.1. Příprava vzorků
Poznámka:
Pro účely detekce latentních populací R. solanacearum se doporučuje
testovat smíšené vzorky. Postup lze vhodně použít na smíšené vzorky o počtu až 200
stonkových částí. Provádí-li se průzkum, měl by být založen na statisticky reprezentativním
vzorku zkoumané rostlinné populace.
3.2.1.1. Do uzavřené sterilní nádoby se uloží 1- 2 cm dlouhé kousky stonků
podle následujícího postupu vzorkování:
Rajčatové sazenice z předpěstírny: Čistým dezinfikovaným nožem se odebere
kousek dlouhý 1 cm z paty každého stonku hned nad úrovní země.
Rostliny z pole nebo skleníku: Čistým dezinfikovaným nožem se odebere nejspodnější
postranní výhonek z každé rostliny uříznutý hned nad spojením s hlavním stonkem.
Odebere se nejspodnější, 1 cm dlouhý díl z každého výhonku. Jiné hostitelské rostliny:
Čistým dezinfikovaným nožem nebo zahradnickými nůžkami se odebere 1 cm dlouhý díl
z paty každého stonku hned nad úrovní země. U lilku potměchuti (S. dulcamara) nebo
jiných hostitelských rostlin rostoucích ve vodě se odeberou 1-2 cm díly ze stonku
pod vodou nebo oddenků s vodními kořeny.
Při vzorkování určité oblasti se doporučuje testovat statisticky representativní
vzorek o počtu nejméně 10 rostlin na 1 vzorkovací místo pro každou potenciální plevelnou
hostitelskou rostlinu. Detekce patogenu bude nejspolehlivější v období pozdního jara,
léta a podzimu, ačkoli přirozenou infekci je možné zjistit po celý rok u víceleté
Solanum dulcamara rostoucí ve vodních tocích. Mezi známé hostitelské rostliny patří
plevelné rostliny bramboru, Solanum dulcamara, S. nigrum, Datura stramonium a další
zástupci čeledi Solanaceae. Dalšími hostitelskými rostlinami jsou rostliny rodu Pelargonium
spp. a Portulaca oleracea. Některé evropské plevelné druhy, které mohou hostit populace
R. solanacearum biovar 2/Race 3 v kořenech a/nebo oddencích za specifických podmínek,
zahrnují Atriplex hastata, Bidens pilosa, Cerastium glomeratum, Chenopodium album,
Eupatorium cannabinum, Galinsoga parviflora, Ranunculus scleratus, Rorippa spp, Rumex
spp., Silene alba, S. nutans., Tussilago farfarra a Urtica dioica.
Poznámka:
V této fázi je možné provést vizuální šetření vnitřních příznaků
(zbarvení cév nebo bakteriální sliz). Jakýkoli díl stonku s příznaky se dá stranou
a testuje se odděleně (viz. oddíl 2.).
3.2.1.2. Části stonku se krátce dezinfikují 70% etanolem a ihned vysuší
savým papírem. Potom se zpracují části stonku jedním z následujících postupů:
a) zalijí se části dostatečným množstvím (přibližně 40 ml) extrakčního
pufru (dodatek 4) a třepou na rotační třepačce (50-100 ot./min) 4 hodiny při teplotě
do 24oC nebo 16-24 hodin chlazené,
nebo
b) se ihned zpracují drcením v pevném maceračním sáčku (např. Stomacher
nebo Bioreba) s přiměřeným množstvím extrakčního pufru (dodatek 4) pomocí gumového
tloučku nebo vhodného mlecího zařízení (např. Homex). Není-li to možné, uloží se
části stonků v chladu nejdéle na 72 hodin nebo při pokojové teplotě (18 - 25oC) na
24 hodin.
3.2.1.3. Po usazení, které má trvat 15 minut, se dekantuje supernatant.
3.2.1.4. Další pročišťování extraktu nebo zahušťování bakteriální frakce
obvykle není třeba, ale lze je provést filtrací a/nebo odstřeďováním popsaným v oddíle
3.1.1.3. až 3.1.1.5.
3.2.1.5. Čistý nebo koncentrovaný vzorkový extrakt se rozdělí na dva stejné
díly. Jeden díl se testuje při teplotě 4-10 st. C a druhý díl se skladuje v 10-25%
(v/v) sterilním glycerolu při teplotě -16 až -24 st. C (týdny) nebo -68 až -86oC
(měsíce) pro případné další testování.
3.2.2. Testování
Postupové diagramy a popisy testů a optimalizovaných protokolů jsou v příslušných
dodatcích:
Selektivní izolace (viz. oddíl 6.1.4.)
Test IF (viz. oddíl 6.1.5.)
Testy
PCR (viz. oddíl 6.1.6.)
Test FISH (viz. oddíl 6.1.7.)
Testy ELISA (viz. oddíl 6.1.8.)
Biotest
(viz. oddíl 6.1.9.)
4. Detekce a identifikace původce hnědé hniloby ve vodě
4.1 Postupový diagram pro detekci a identifikaci R. solanacearum ve vodě

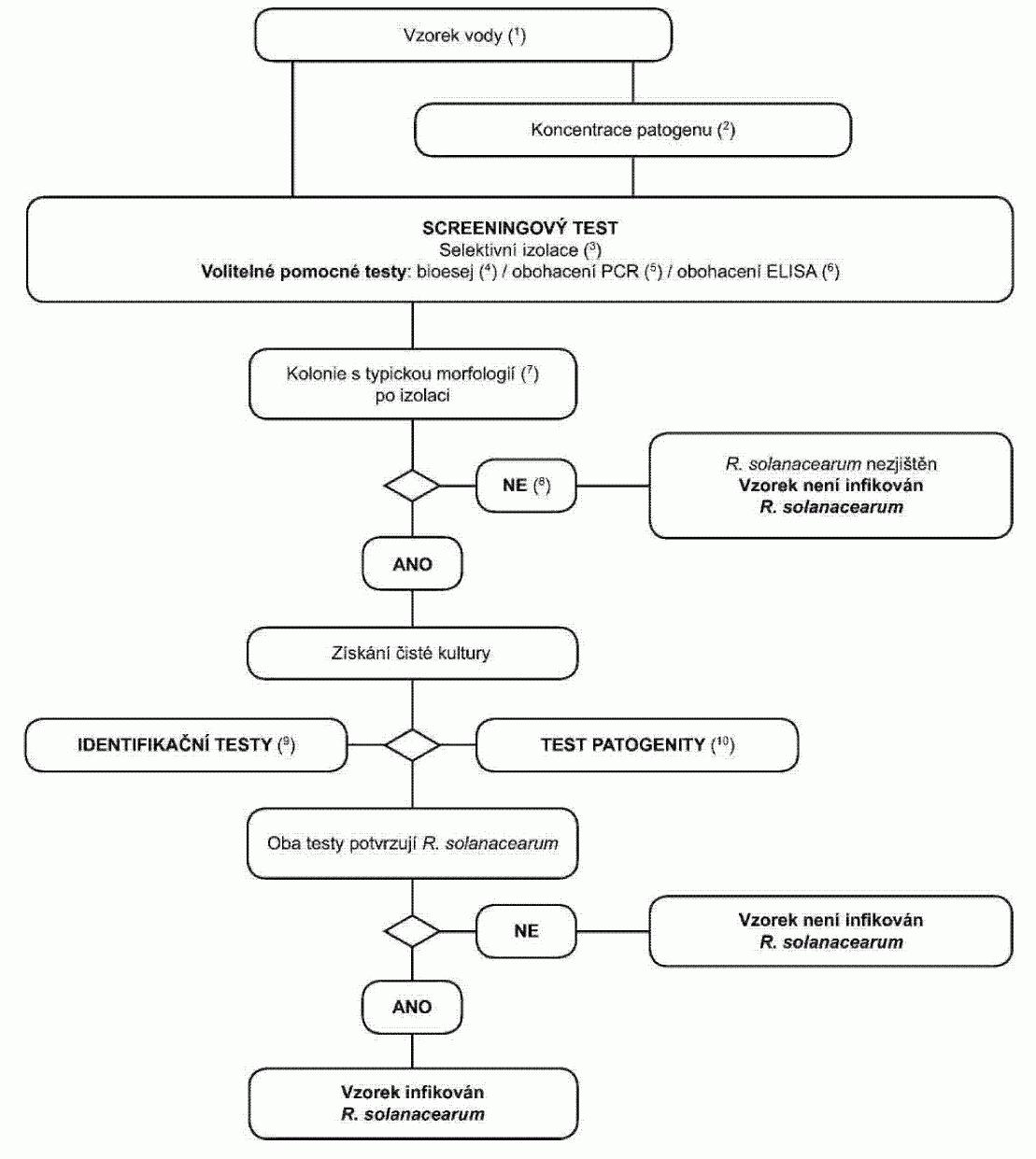
-----
(1) Viz. oddíl 4.2.1., kde jsou uvedeny doporučené postupy vzorkování.
(2) Metody koncentrace patogenu jsou popsány v oddíle 4.2.1. Koncentrace
zvětšuje populaci jak patogenu, tak konkurenčních saprofytických bakterií a doporučuje
se jen tehdy, jestliže nebrání izolačnímu testu.
(3) Selektivní izolační test je popsán v oddíle 6.1.4.
(4) Biotest je popsán v oddíle 6.1.9.
(5) Metody obohacovacího testu PCR jsou popsány v oddíle 6.1.4.2. a 6.1.6.
(6) Metody obohacovacího testu ELISA jsou popsány v oddíle 6.1.4.2. a
6.1.8.
(7) Typická morfologie kolonie je popsána v oddíle 2.3.d.
(8) Kultivace může selhat z důvodu konkurence nebo inhibice saprofytickými
bakteriemi. Je-li podezření, že velká saprofytická populace ovlivní spolehlivost
izolace, opakují se izolační testy po zředění vzorku sterilní vodou.
(9) Spolehlivá identifikace podezřelých čistých kultur R. solanacearum
se dosáhne pomocí testů popsaných v oddíle 6.2.
(10) Test patogenity je popsán v oddíle 6.3.
4.2. Metody detekce a identifikace R. solanacearum ve vodě
Princip
Validované schéma detekce popsané v tomto oddíle je použitelné k
detekci patogenu ve vzorcích povrchové vody a rovněž může být použito k testování
vzorků odpadní vody ze zpracování brambor a odpadní vody. Očekávaná citlivost detekce
se však liší v závislosti na substrátu. Citlivost testu selektivní izolace je ovlivňována
populacemi konkurenčních saprofytických bakterií, kterých je obecně mnohem více v
odpadní vodě ze zpracování brambor a odpadní vodě než v povrchové vodě. Zatímco níže
uvedené schéma předpokládá detekci asi 10
3
buněk/litr povrchové vody, citlivost detekce
v odpadní vodě ze zpracování brambor a odpadní vodě bude pravděpodobně podstatně
nižší. Z tohoto důvodu se doporučuje testovat odpadní vodu po provedení nějakého
čistícího postupu (např. sedimentace nebo filtrace), při nichž dojde ke snížení saprofytických
bakteriálních populací. Omezení citlivosti testovacího schématu by mělo být bráno
v úvahu při hodnocení spolehlivosti v případě získání negativních výsledků.Vzhledem
k tomu, že se toto schéma používá úspěšně k mapování výskytu nebo nepřítomnosti patogenu
v povrchové vodě, je třeba si uvědomit jeho omezení při použití k podobnému mapování
v odpadní vodě ze zpracování brambor a v odpadní vodě. 4.2.1. Příprava vzorků
Poznámka:
- Zjištění R. solanacearum v povrchové vodě je nejspolehlivější
v období pozdního jara, léta a podzimu, kdy teplota vody překračuje 15 st. C.
- Opakované
vzorkování v různé době během výše uvedených období na určených vzorkovacích místech
zvýší spolehlivost zjištění snížením vlivů výkyvů povětrnostních podmínek.
- Vlivem
silných dešťů a geografie vodního toku může dojít ke značnému zředění a tím i zastření
výskytu patogenu.
- Vzorky vody se odebírají v blízkosti hostitelských rostlin, pokud
jsou přítomny.
4.2.1.1. Na vybraných vzorkovacích místech se odeberou vzorky vody do sterilních
zkumavek nebo lahviček na jedno použití, pokud možno 30 cm pod hladinou a 2 m od
břehu. Při vzorkování odpadní vody ze zpracování brambor a odpadní vody se odeberou
vzorky z místa výtoku odpadní vody. Doporučená velikost vzorku je 500 ml. Je-li upřednostňována
menší velikost, doporučuje se odebrat vzorky nejméně 3krát pro každé vzorkovací místo,
z nichž každý bude obsahovat 2 opakované dílčí vzorky o velikosti nejméně 30 ml.
Při intenzivním mapování se vyberou nejméně 3 vzorkovací místa na každé 3 km vodního
toku a vzorkují se rovněž přítoky.
4.2.1.2. Vzorky se přepravují v chladu a temnu (4-10 st. C) a testují do
24 hodin.
4.2.1.3. Je-li potřeba, může být provedena koncentrace bakteriální frakce
pomocí následujících metod:
a) Odstřeďují se 30-50 ml dílčí vzorky při 10 000 g po dobu 10 minut (nebo
7000 g po dobu 15 minut), pokud možno při teplotě 4-10 st. C, slije se kapalina nad
usazeninou a resuspenduje se peleta v 1 ml pufru (dodatek 4).
b) Provede se filtrace přes membránu (minimální velikost pórů 0,45 µm)
s následným promytím filtru v 5-10 ml peletového pufru a zachycením filtrátu. Tato
metoda je vhodná pro velké objemy vody, které obsahují malý počet saprofytů.
Koncentrace se obvykle nedoporučuje u vzorků vody ze zpracování brambor nebo
u vzorků odpadní vody, protože zvýšená populace konkurenčních saprofytických bakterií
inhibuje detekci Ralstonia solanacearum.
4.2.2. Testování
Viz. postupový diagram a popis testů v příslušných dodatcích.
5. Detekce a identifikace původce hnědé hniloby v půdě
5.1. Postupový diagram pro detekci a identifikaci R. solanacearum v půdě

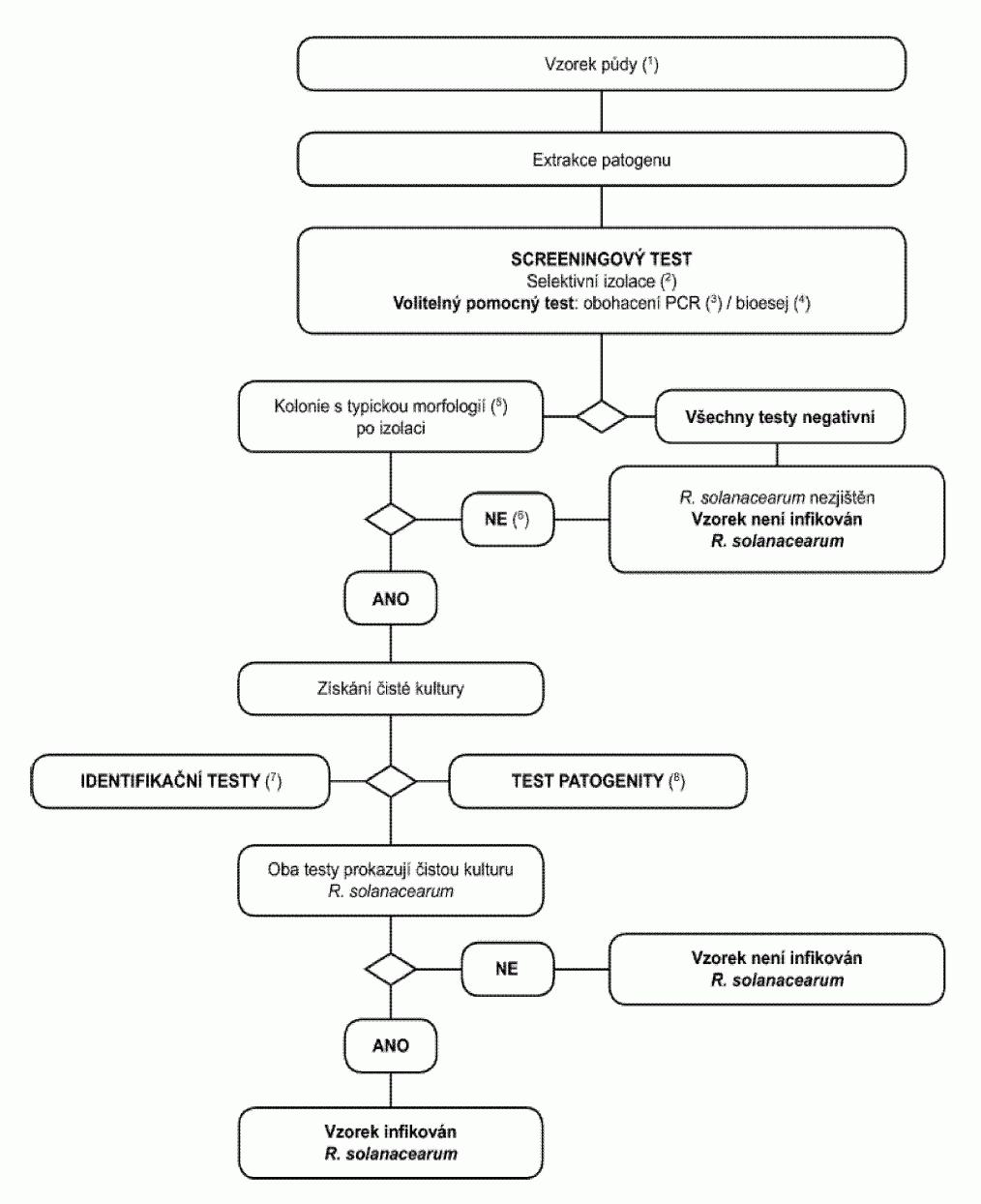
-----
(1) Viz. oddíl 5.2.1., kde jsou uvedeny doporučené postupy vzorkování
(2) Selektivní izolační test je popsán v oddíle 6.1.4.
(3) Obohacovací PCR testy jsou popsány v oddíle 6.1.4.2. a 6.1.6.
(4) Biotest je popsán v oddíle 6.1.9.
(5) Typická morfologie kolonie je popsána v oddíle 2.3.d.
(6) Kultivace může selhat z důvodů konkurence nebo inhibice saprofytickými
bakteriemi. Je-li podezření na vliv saprofytické populace na spolehlivost izolace,
opakují se testy selektivní izolace po dalším zředění vzorku.
(7) Spolehlivá identifikace podezřelých čistých kultur R. solanacearum
se dosáhne pomocí testů popsaných v oddíle 6.2.
(8) Test patogenity je popsán v oddíle 6.3.
5.2. Metody pro detekci a identifikaci R. solanacearum v půdě
Principy
Platné detekční schéma popsané v tomto oddíle je použitelné pro
detekci patogenu ve vzorcích půdy, ale může být rovněž použito k testování vzorků
pevného odpadu při zpracování brambor nebo kalu z odpadní vody. Je však nutné poznamenat,
že tyto metody nejsou dostatečně citlivé, aby zaručovaly detekci nízkých nebo nepravidelně
rozptýlených populací Ralstonia solanacearum, které se mohou vyskytovat v přirozeně
infikovaných vzorcích těchto substrátů.
Při hodnocení spolehlivosti všech získaných
negativních výsledků a také při sestavování přehledů určujících přítomnost patogenu
v půdě nebo kalu je třeba vzít v úvahu omezenou citlivost tohoto testovacího schématu.
Nejspolehlivějším testem přítomnosti patogenu v polní půdě je vypěstování náchylné
hostitelské rostliny a její sledování, zda není infikována, ale i při použití této
metody unikne nízký stupeň zamoření pozornosti.
5.2.1. Příprava vzorků
5.2.1.1. Vzorkování polní půdy by se mělo řídit standardními principy používanými
pro vzorkování hlístů. Na jeden vzorek se shromáždí 0,5 až 1 kg půdy ze 60 míst na
ploše 0,3 ha z hloubky 10-20 cm. Je-li podezření na výskyt patogenu, zvyší se počet
sběrných míst na 120 z plochy 0,3 ha. Před testováním se uchovávají vzorky při teplotě
12-15 st. C. Kal ze zpracování brambor a kanalizace se navzorkuje shromážděním celkem
1 kg z míst reprezentujících celkový objem testovaného kalu. Před testováním se každý
vzorek dobře promíchá.
5.2.1.2. Dílčí vzorky 10-25 g půdy nebo kalu se rozptýlí rotační třepačkou
(250 ot./min) v 60-150 ml extrakčního pufru (dodatek 4) po dobu 2 hodin. Je-li třeba,
přidání 0,02% sterilního Tween-20 a 10-20 g sterilních kamínků může napomoci disperzi.
5.2.1.3. Během testování se udržuje suspenze v teplotě 4 st. C.
5.2.2. Testování
Postupový diagram a popis testů jsou v příslušném dodatku.
6. Optimalizované protokoly pro detekci a identifikaci R. solanacearum
6.1. Diagnostické a detekční testy
6.1.1. Test na výtok slizu ze stonku
Přítomnost R. solanacearum ve stoncích
vadnoucích rostlin bramboru, rajčete nebo jiných hostitelských rostlin může ukázat
následující jednoduchý test pravděpodobného výskytu: uřízne se stonek právě nad úrovní
země. Řez stonku se ponoří do zkumavky s čistou vodou. Sleduje,se, zda se po několika
minutách objeví charakteristická samovolně proudící vlákna bakteriálního slizu z
přeříznutých cévních svazků.
6.1.2. Test na přítomnost granulí poly-beta- hydroxybutyrátu
1. Připraví se roztěr bakteriálního slizu z infikovaného pletiva nebo ze
48 hodinové kultury na živné půdě YPGA nebo SPA (dodatek 2) na mikroskopické sklíčko.
2. Připraví se pozitivní kontrolní roztěr kmenu biovaru 2 R. solanacearum
a případně negativní kontrolní roztěr o známém PHB negativním sp.
3. Roztěry se nechají uschnout a spodní povrch každého sklíčka se rychle
protáhne nad plamenem, aby se roztěry fixovaly.
4. Preparáty se obarví buď nilskou modří nebo súdánskou černí a provede se
mikroskopické pozorování podle níže uvedeného popisu:
Test nilskou modří:
a) Obě sklíčka se zakápnou 1% vodným roztokem nilské modři a nechají se
inkubovat 10 minut při teplotě 55 st. C.
b) Odstraní se barvící roztok. Krátce se opláchne jemně tekoucí vodou z
kohoutku. Přebytečná voda se odsaje savým papírem.
c) Roztěr se zakápne 8% vodním roztokem kyseliny octové a nechá se inkubovat
1 minutu při pokojové teplotě.
d) Krátce se opláchne jemně tekoucí vodou z kohoutku.
Přebytečná voda se
odsaje savým papírem.
e) Znovu se navlhčí kapkou vody a přiloží se krycí sklíčko.
f) Zkoumá se obarvený roztěr epifluorescenčním mikroskopem při 450 nm pod
olejovou nebo vodní imersí se zvětšením 600x až 1000x s použitím objektivu pro olejovou
nebo vodní imersi.
g) Pozoruje se, zda se objeví na PHB granulích jasně oranžová fluorescence.
Také se pozoruje v procházejícím normálním světle, zda jsou granule intracelulární
a zda je morfologie buněk typická pro R. solanacearum.
Test súdánskou černí:
a) Zakápnou se všechna sklíčka 0,3% roztokem súdánské černi B v 70% etanolu
a inkubují se 10 minut při pokojové teplotě.
b) Odstraní se barvící roztok a krátce se opláchne jemně tekoucí vodou
z kohoutku a odsaje se přebytečná voda savým papírem.
c) Sklíčka se ponoří krátce do xylolu a vysají se savým papírem. Upozornění
: Xylol je zdraví škodlivý, je třeba dodržet nezbytná bezpečnostní opatření a pracovat
v digestoři.
d) Sklíčka se zakápnou 0,5% (w/v) vodným roztokem safraninu a nechají 10
vteřin inkubovat při pokojové teplotě.
Upozornění: safranin je zdraví škodlivý, je třeba dodržet nezbytná bezpečnostní
opatření a pracovat v digestoři.
e) Sklíčka se opláchnou jemně tekoucí vodou z kohoutku, odsaje se přebytečná
voda savým papírem a přiloží se krycí sklíčko.
f) Zkoumá se obarvený roztěr mikroskopem s procházejícím světlem pod olejovou
imersí a při zvětšení 1 000x.
g) Hledá se modročerné zbarvení PHB granulí v buňkách R. solanacearum s
růžově zbarvenými stěnami buněk.
6.1.3. Sérologické aglutinační testy
Aglutinace buněk R. solanacearum v bakteriálním slizu nebo v extraktech
z pletiv s příznaky je nejlépe pozorovatelná pomocí validovaných protilátek (viz.
dodatek 3) označených příslušnými obarvenými značkovači, např. červenými buňkami
Staphylococcus aureus nebo obarvenými latexovými částicemi. Při použití komerčně
dostupného vybavení (viz. dodatek 3) se postupuje podle instrukcí výrobce. Jinak
je postup následující:
a) Smíchají se kapky suspenze označené protilátky a bakteriálního slizu
(přibližně 5 µl každé látky) na okénku testovacího víceokénkového sklíčka.
b) Připraví se pozitivní a negativní kontrolní vzorky použitím suspenzí
biovaru 2 R. solanacearum a heterologního kmenu.
c) Pozoruje se, zda se v pozitivních vzorcích po jemném míchání po dobu
15 sekund objeví aglutinace.
6.1.4. Selektivní izolace
6.1.4.1. Očkování na živnou půdu
Poznámka:
Než se použije tato metoda poprvé, provede se předběžný test k
zajištění reprodukovatelné detekce 10
3
až 104
jednotek tvořících kolonie R. solanacearum
na 1 ml přidaných do extraktů ze vzorků, které byly předtím testovány s negativním
výsledkem.Použije se řádně validovaná selektivní živná půda, např. SMSA (ve znění
Elphinstone et al., 1996; viz. dodatek 2).
Rozlišení R. solanacearum od jiných bakterií
schopných na živné půdě růst vyžaduje velkou pozornost. Dále, kolonie R. solanacearum
mohou mít atypickou morfologii, jestliže Petriho misky jsou přeplněny nebo jsou rovněž
přítomny antagonistické bakterie. Je-li podezření na konkurenci nebo inhibici, měl
by být vzorek otestován znovu pomocí jiného testu.
Při použití čerstvě připravených
extraktů ze vzorků lez očekávat, že citlivost detekce touto metodou bude velmi vysoká.
Metoda je však použitelná i pro extrakty, které byly uchovány v glycerolu při teplotě
od -68 do -86 st. C.
Pozitivní kontroly se připraví jako desetinné ředění ze suspenze 106 cfu/ml
virulentního kmene biovaru 2 R. solanacearum (např. NCPPB 4156 = PD 2762= CFBP 3857).
K vyloučení možné kontaminace se připraví pozitivní kontrolní vzorky zcela odděleně
od vzorků k testování. U každé nově připravené šarže selektivní živné půdy by měla
být před jejím použitím k rutinnímu testování vzorků nejdříve otestována její vhodnost
pro kultivaci patogenu. Kontrolní materiál se testuje týmž způsobem jako vzorek (vzorky).
1. Provede se ředění s cílem zajistit, aby všechny populace saprofytických
bakterií byly vyloučeny. Nanese se 50- 100 µl extraktu vzorku na 1 misku pro každé
ředění.
2. Misky se nechají inkubovat při teplotě 28 st. C. Po 48 hodinách se zkontroluje
jejich stav a pak se provádí kontrola denně po 6 dní. Typické kolonie R. solanacearum
na živné půdě SMSA jsou mléčně bílé, ploché, nepravidelné a fluidní a po 3 dnech
inkubace se rozvine růžově až krvavě červené zbarvení ve středu s vnitřními pruhy
nebo spirálami.
Poznámka:
Někdy se na tomto médiu tvoří atypické kolonie R. solanacearum.
Mohou být malé, kruhové, zcela červené a nefluidní nebo jen částečně fluidní, a proto
obtížně rozeznatelné od saprofytických bakterií tvořících kolonie.
3. Předpokládané kolonie R. solanacearum se rozetřou nebo očkují metodou
zřeďování na universální živnou půdu, aby se získaly izolované kolonie (viz. dodatek
2).
4. Kultury se uchovávají krátkodobě ve sterilní vodě (pH 6-8, bez chlóru)
při pokojové teplotě v temnu nebo dlouhodobě ve vhodném ochranném médiu při teplotě
-68 až -86oC nebo lyofilizované.
5. Provede se identifikace podezřelých kultur (viz. oddíl 6.2.) a test patogenity
(viz oddíl 6.3).
Interpretace výsledků testu očkováním na selektivní média. Test očkování
na selektivní média je negativní, jestliže po 6 dnech nejsou zpozorovány žádné bakterie
nebo nejsou nalezeny žádné podezřelé kolonie typické pro R. solanacearum , za předpokladu,
že nedošlo k inhibici jinými bakteriemi a v kontrolních vzorcích jsou nalezeny typické
kolonie R. solanacearum.
Test očkování na selektivní média je pozitivní, jestliže jsou izolovány podezřelé
kolonie R. solanacearum.
6.1.4.2. Obohacovací testy
Použije se validované médium pro obohacení, např. modifikovaný bujón Wilbrink
(viz dodatek 2).
Tento postup lze použít pro selektivní zvětšení populací R. solanacearum
v extraktech ze vzorků a zvýšení citlivosti detekce. Tímto způsobem dojde i ke zředění
potenciálních inhibitorů PCR testu (1:100). Obohacení R. solanacearum však může být
neúspěšné z důvodů konkurence nebo antagonismu saprofytických organismů, které bývají
často současně také obohaceny. Z tohoto důvodu může být izolace R. solanacearum z
obohacené kultury obtížná. Kromě toho, protože může dojít k rozvoji populací sérologicky
příbuzných saprofytů, se pro test ELISA doporučuje použít místo polyklonálních protilátek
specifické monoklonální protilátky.
1. U obohacovacího testu PCR se přenese 100 ul extraktu ze vzorku do 10 ml
obohaceného bujónu (dodatek 2), který se předem připraví do sterilních zkumavek nebo
baněk. U obohacovacího testu ELISA může být použit větší podíl extraktu ze vzorku
do bujónu (např. 100 ul v 1,0 ml obohacené živné půdy).
2. Inkubuje se 72 hodin při teplotě 27 až 30 °C v třepané nebo statické kultuře
s uvolněnými uzávěry, které umožní provzdušňování.
3. Před zahájením testů ELISA nebo PCR se obsah dobře promíchá.
4. S obohaceným bujónem se zachází týmž způsobem jako se vzorky ve výše uvedených
testech.
Poznámka:
Očekává-li se inhibice obohacení R. solanacearum z důvodu vysoké
koncentrace konkurenčních saprofytických bakterií, lze docílit lepších výsledků obohacením
extraktů ze vzorků před jakýmkoli odstřeďováním nebo jinými postupy zvyšování koncentrace.
6.1.5. Test IF
Postup
Použití testu IF jako hlavního screeningového testu se doporučuje
kvůli dokázané spolehlivosti v dosažení požadovaných prahů.
Použije-li se test IF jako hlavní a výsledek IF testu je pozitivní, musí
být jako druhý screeningový test použit test izolace, PCR nebo FISH. Jestliže je
test IF použit jako druhý screeningový test a je pozitivní, je nutné provést další
testování podle postupového diagramu, aby byla analýza úplná.
Poznámka:
Použije se validovaný zdroj protilátek R. solanacearum. Doporučuje
se určit titr pro každou novou šarži protilátek. Titr je definován jako nejvyšší
ředění, při kterém dojde k optimální reakci při testování suspenze obsahující 10
5
až 106
buněk/ml homologického kmene R. solanacearum a použitím vhodného konjugátu
fluorescenčního isothiokyanatanu (FITC) podle doporučení výrobce. Validovaná polyklonální
antiséra mají všechna titr IF nejméně 1:2 000. Během testování by měly být použity
protilátky v pracovních ředěních blízkých nebo stejných jako titr.Test by měl být proveden na čerstvě připravených extraktech ze vzorků. Jestliže
je to nutné, může být úspěšně proveden na vzorcích uchovaných při teplotě - 68 až
- 86 °C v glycerolu. Glycerol může být ze vzorku odstraněn přidáním 1 ml pufru (dodatek
4), 15minutovým odstřeďováním při 7000 g a resuspenzí ve stejném množství peletového
pufru. Často to není potřeba, zvláště pokud jsou vzorky fixovány na sklíčka plamenem.
Připraví se oddělená pozitivní kontrolní sklíčka s homologickým kmenem nebo jiným
referenčním kmenem R. solanacearum suspendovaným v bramborovém extraktu, jak je uvedeno
v dodatku 3 B, a nepovinně v pufru.
Pletivo infikované přirozenou cestou (uchovávané lyofilizací nebo zmrazením
při teplotě - 16 až 24 °C) by se mělo podle možností použít jako paralelní kontrola
na stejném podložním sklíčku.
Jako negativní kontroly se použijí alikvotní díly extraktů ze vzorků, které
byly předtím testovány s negativním výsledkem.
Standardizované materiály k pozitivní a negativní kontrole, které lze použít
pro tento test, jsou uvedeny v dodatku 3. Použijí se mikroskopická sklíčka s více
okénky, pokud možno s 10 okénky s průměrem nejméně 6 mm.
Test kontrolního materiálu se provede týmž způsobem jako test vzorků.
6.1.5.1. Připraví se sklíčka k testování jedním z následujících postupů:
i) Pro pelety s relativně nízkým množstvím sedimentu škrobu:
Napipetuje
se odměřené standardní množství (15 µl je vhodné pro průměr okénka 6 mm - u většího
okénka je množství větší v příslušném poměru) resuspendované bramborové pelety v
ředění 1/100 do prvního okénka. Následně se napipetuje stejné množství nezředěné
pelety (1/1) do zbývajících okének v řadě. Druhá řada může být použita jako duplikát
nebo pro druhý vzorek, jak ukazuje obr. 1.
ii) Pro jiné pelety:
Připraví se desetinná ředění (1/10, 1/100) resuspendované
pelety v peletovém pufru. Napipetuje se odměřené standardní množství (15 µl je vhodné
pro průměr okénka 6 mm - u většího okénka je množství větší v příslušném poměru)
resuspendované pelety pro každé ředění do řady okének. Druhá řada může být použita
jako rezervní nebo pro druhý vzorek, jak ukazuje obr.2.
6.1.5.2. Vysuší se kapičky při okolní teplotě nebo zahřátím na teplotu 40
až 45 °C. Bakteriální buňky se fixují na sklíčko buď zahřátím (15 minut při teplotě
60 °C), plamenem, 95 % etanolem nebo podle konkrétních instrukcí od dodavatele protilátek.
Pokud je potřeba, fixovaná sklíčka je možné skladovat ve zmrazeném stavu v suchém
boxu po nezbytně krátkou dobu (maximálně 3 měsíce) před dalším testováním.
6.1.5.3. Postup IF
i) Podle přípravy sklíčka na test v odst. 5.1.i): Připraví se sada dvojnásobných
zředění. První jamka by měla mít 1/2 titru (T/2), ostatní 1/4 titru (T/4), 1/2 titru
(T/2), titr (T) a dvojnásobek titru (2T).
ii) Podle přípravy sklíčka na test v odst. 5.1.ii): Připraví se pracovní
ředění (PŘ) protilátek v pufru IF. Pracovní ředění ovlivňuje přesnost.

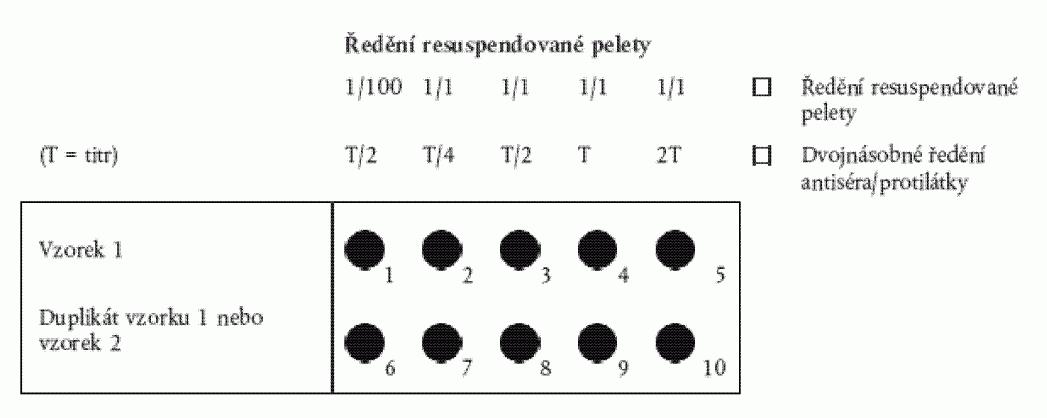
Obrázek 1. Příprava sklíčka na test podle odst. 6.1.5.1.i) a 6.1.5.3.i)

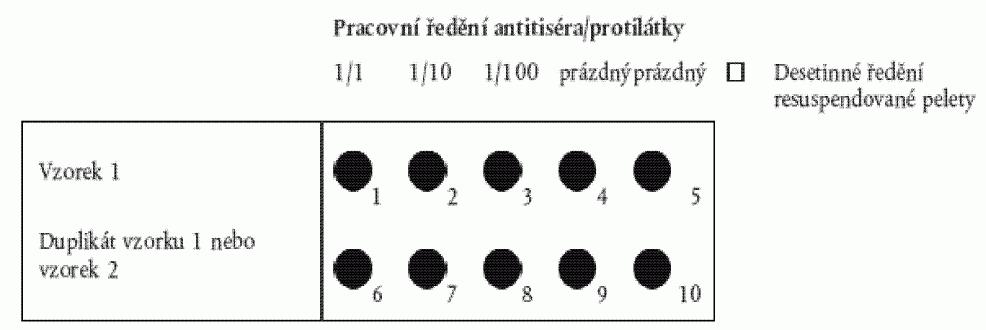
Obrázek 2. Příprava sklíčka na test podle odst. 6.1.5.1.ii) a 6.1.5.3.ii)
1. Podložní sklíčka se uspořádají na navlhčený savý papír. Každé testovací
okénko se pokryje kompletně ředěním protilátek. Množství protilátky na každém okénku
musí být nejméně stejné jako množství použitého extraktu.
Pokud nejsou k dispozici
konkrétní pokyny od dodavatele protilátek, postupuje se následovně:
2. Podložní sklíčka se inkubují na vlhkém papíře přikrytá po dobu 30 minut
při pokojové teplotě (18-25 st. C).
3. Setřesou se kapky ze všech podložních sklíček a tato se pečlivě opláchnou
pufrem IF. Umyjí se ponořením po dobu 5 minut v pufru IF Tween (dodatek 4) a následně
v pufru IF. Je třeba zabránit vzniku aerosolu nebo přenosu kapiček, které by mohly
způsobit vzájemnou kontaminaci. Pečlivě se odstraní přebytečná vlhkost jemným osušením.
4. Sklíčka se umístí na vlhký papír. Testovací okénka se pokryjí ředěním
konjugátu FITC, kterým se stanovuje titr. Množství konjugátu naneseného do okének
musí být stejné jako množství použité protilátky.
5. Sklíčka se inkubují zakrytá na vlhkém papíru po dobu 30 minut při pokojové
teplotě (18-25 st. C).
6. Setřesou se kapky konjugátu ze sklíčka. Sklíčko se opláchne a umyje jako
předtím (3.). Opatrně se odstraní přebytečná vlhkost.
7. Napipetuje se 5-10 ul 0,1M fosfátového pufru s glycerolem (dodatek 4)
nebo komerční krycí tekutiny do každého okénka a přiloží se krycí sklíčko.
6.1.5.4. Vyhodnocení IF testu
1. Prohlížejí se testovací sklíčka epifluorescenčním mikroskopem s filtry
vhodnými pro excitaci FITC pod olejovou nebo vodní imersí při zvětšení 500x až 1
000x. Zkoumají se okénka ve dvou navzájem kolmých průměrech a kolem obvodu. U vzorků
s žádnými nebo malým počtem buněk se zkoumá nejméně 40 polí mikroskopu.
Nejdřív se
zkontroluje pozitivní kontrolní vzorek. Buňky musí být jasně fluoreskující a zcela
obarvené v určeném titru protilátky nebo pracovním ředění. Pokud je barevnost odchylná,
musí být test IF opakován (odst. 6.1.5).
2. Pozorují se jasně fluoreskující buňky s charakteristickou morfologií R.
solanacearum v testovacích okéncích sklíčka. Intenzita fluorescence musí být při
porovnání s pozitivním kontrolním kmenem ve stejném ředění protilátky stejná nebo
lepší. Buňky s neúplným zbarvením nebo slabou fluorescencí nelze brát v úvahu.
Při
podezření z jakékoli kontaminace musí být test zopakován. To se může stát, když všechna
sklíčka ve skupině vykazují pozitivní buňky díky kontaminaci pufru nebo při zjištění
pozitivních buněk (mimo okénka sklíček) na povrchu sklíček.
3. Existuje několik problémů podstatných pro přesnost imunofluorescenčního
testu. V peletách z pupkové části bramboru a částí stonku se mohou vyskytnout doprovodné
populace fluoreskujících buněk s atypickou morfologií a křížově reagující saprofytické
bakterie s velikostí a morfologií podobnou R. solanacearum.
4. Berou se v úvahu pouze fluoreskující buňky s typickou velikostí a morfologií
v titru nebo pracovním ředění protilátek podle oddílu 6.1.5.3.
5. Interpretace výsledků testu IF:
i) Při zjištění jasně fluoreskujících buněk s typickou morfologií se odhadne
průměrný počet typických buněk v 1 mikroskopovém poli a vypočítá počet typických
buněk na 1 ml resuspendované pelety (dodatek 5).
Výsledek IF je pozitivní u vzorků,
kde je počet typických buněk na 1 ml resuspendované pelety nejméně 5 x 10
3
. Vzorek
je považován za potenciálně infikovaný a je povinné další testování.ii) Výsledek IF testu je negativní pro vzorky, které obsahují méně než
5 x 10
3
buněk na 1 ml resuspendované pelety a vzorek se považuje za negativní. Další
testování není nutné. 6.1.6. Testy PCR
Principy
Použije-li se test PCR jako hlavní screeningový test a výsledek
je pozitivní, musí být povinně proveden druhý screeningový test, a sice izolace nebo
IF. Jestliže je PCR použita jako druhý screeningový test a je pozitivní, je nutné
provést další testování podle postupového diagramu, aby byla analýza úplná.
Využití této metody v celém rozsahu jako hlavního screeningového testu se
doporučuje jen tehdy, je-li požadována specializovaná expertíza.
Poznámka:
Předběžné testování touto metodou by mělo umožnit reprodukovatelné
zjištění 10
3
až 104
buněk R. solanacearum na 1 ml přidaný do vzorků extraktů, které
byly předtím testovány jako negativní. Dosažení maximální citlivosti a přesnosti
ve všech laboratořích může vyžadovat optimalizační pokusy. Použijí se schválená činidla
a protokoly PCR (viz dodatek 6). Pokud možno, zvolí se metoda s interní kontrolou.Je třeba použít vhodná bezpečnostní opatření k zabránění kontaminace vzorku
cílovou DNA. Test PCR by měl být prováděn zkušenými laboranty v laboratořích specializovaných
na molekulární biologii, aby se minimalizovala možnost kontaminace cílovou DNA.
S negativními kontrolami (v průběhu extrakce DNA a PCR) by se mělo vždy zacházet
jako s konečnými vzorky, aby bylo jasné, jestli došlo k přenosu DNA.
PCR test by měl zahrnovat následující negativní kontroly:
- extraktu ze vzorku,
který byl předtím testován na R. solanacearum s negativním výsledkem,
- kontroly pufru
používaného pro extrakci bakterií a DNA ze vzorku,
- reakční směs PCR.
Měly by být
zahrnuty následující pozitivní kontroly:
- alikvotní části resuspendovaných pelet,
do kterých byla přidána R. solanacearum (přípravu viz dodatek 3 B);
- suspenze 106
buněk na 1 ml R. solanacearum ve vodě z virulentního izolátu (např. NCPPB 4156 =
PD 2762 = CFBP 3857; viz dodatek 3 B);
- pokud možno, při PCR použít také DNA extrahovanou
z pozitivních kontrolních vzorků.
Aby se zabránilo případné kontaminaci, připravují se pozitivní kontroly v
prostředí odděleném od vzorků, které budou testovány.
Extrakty ze vzorků by měly být pokud možno bez zeminy. V případě použití
PCR testu je potřeba připravit extrakty z umytých brambor.
Standardizované materiály k pozitivní a negativní kontrole, které lze použít
pro tento test, jsou uvedeny v dodatku 3.
6.1.6.1. Metody purifikace DNA
Používají se výše popsané pozitivní a negativní kontrolní vzorky (viz. dodatek
3).
Test kontrolního materiálu se provede týmž způsobem jako test vzorků.
K purifikaci cílové DNA z komplexních substrátů vzorků jsou k dispozici různé
metody odstraňující inhibitory PCR a jiných enzymových reakcí a koncentrující cílovou
DNA ve vzorku. Následující metoda byla optimalizována pro použití se schválenými
metodami PCR uvedenými v dodatku 6.
a) Metoda podle Patrika (2000)
1. Napipetuje se 220 µl lyzátového pufru (100mM NaCl, 10 mM Tris-HCl (pH
8,0), 1 mM EDTA (pH 8,0)) do 1,5 ml Eppendorfovy mikrozkumavky.
2. Přidá se 100 µl extraktu ze vzorku a mikrozkumavka se umístí do termobloku
nebo vodní lázně s teplotou 95 °C na 10 min.
3. Mikrozkumavka se vloží na 5 min. do ledu.
4. Přidá se 80 µl zásobního roztoku lysozymu (50 mg lysozymu/na 1 ml 10 mM
TrisHCl, pH 8,0) a inkubuje při teplotě 37 °C po dobu 30 minut.
5. Přidá se 220 µl roztoku Easy DNA(R) A (Invitrogen), dobře promíchá třepáním
a inkubuje se při teplotě 65 °C po dobu 30 minut.
6. Přidá se 100 µl roztoku Easy DNA(R) B (Invitrogen), důkladně promíchá
třepáním, pokud vzorek nezačne být stejnoměrně viskózní.
7. Přidá se 500 µl chloroformu a promíchá, až se viskozita sníží a směs se
stane homogenní.
8. Pro oddělení fází a vytvoření mezifáze se směs odstřeďuje při 15 000 g
po dobu 20 min. při teplotě 4 °C.
9. Horní fáze se převede do nové Eppendorfovy mikrozkumavky.
10. Přidá se 1 ml 100 % etanolu (- 20 °C), krátce promíchá třepáním a inkubuje
se na ledu po dobu 10 min.
11. Odstřeďuje se při 15 000 g po dobu 20 minut při teplotě 4 °C a odstraní
se etanol z pelety.
12. Přidá se 500 µl 80 % etanolu (- 20 °C) a promíchá se překlápěním mikrozkumavky.
13. Odstřeďuje se při 15 000 g po dobu 10 minut při teplotě 4 °C, zachová
se peleta a odstraní etanol.
14. Peleta se nechá uschnout na vzduchu nebo v DNA odparce.
15. Peleta se resuspenduje v 100 µl sterilní UPW a nechá se stát při pokojové
teplotě nejméně 20 minut.
16. Skladuje se při teplotě - 20 °C až do použití při PCR.
17. Jakákoliv bílá sraženina se odstraní odstředěním. 5 µl supernatantu obsahujícího
DNA se použije pro test PCR.
b) Jiné metody
Jiné metody extrakce DNA, např. Qiagen DNeasy Plant Kit, by se mohly použít,
pokud by se prokázalo, že jsou při purifikaci DNA z kontrolních vzorků obsahujících
10
3
až 104
patogenních buněk na 1 ml stejně efektivní. 6.1.6.2. PCR
1. Připraví se testované vzorky a kontroly pro PCR podle schválených protokolů
(6.1.6). Připraví se desetinásobné ředění vzorku extraktu DNA (1:10 ve sterilní vodě).
2. Připraví se příslušná reakční směs pro PCR v prostředí, ve kterém nehrozí
kontaminace podle zveřejněných protokolů (dodatek 6). Pokud možno, doporučuje se
použít multiplexní protokol PCR, který rovněž zahrnuje interní protokol PCR.
3. Do sterilních PCR mikrozkumavek se přidá 5 µl extraktu DNA na 25 µl PCR
reakce podle protokolů PCR (viz dodatek 6).
4. Přidají se negativní kontrolní vzorky obsahující pouze reakční směs PCR
a přidá se týž zdroj UPW jako byl ten, který byl použit ve směsi PCR místo vzorku.
5. PCR mikrozkumavky se umístí do téhož termocykleru, který byl použit při
počátečním testování, a spustí se vhodně optimalizovaný program PCR (dodatek 6).
6.1.6.3. Analýza produktu PCR
1. Amplikony se rozdělí elektroforézou v agarózovém gelu. Nanese se nejméně
12 µl směsi amplifikované DNA z každého vzorku s 3 µl nanášecího pufru (dodatek 6)
do 2,0 % (w/v) agarózového gelu v Tris-acetát-EDTA (TAE) pufru (dodatek 6) při 5-8
V/cm. Použije se vhodný DNA marker, např. (ladder) 100 bp.
2. Detekují se proužky DNA obarvením v ethidiumbromidu (0,5 mg/l) po dobu
30-60 minut za použití vhodných bezpečnostních opatření pro zacházení s tímto mutagenem.
3. V obarveném a UV (krátké vlnové délky např. 302 nm) prosvíceném gelu se
hledají amplifikované produkty PCR o očekávané velikosti a výsledek se zdokumentuje.
4. U všech nových nálezů/případů se zkontroluje pravost amplikonu PCR provedením
restrikční enzymové analýzy ve zbývajícím vzorku amplifikované DNA inkubací při optimální
teplotě a čase shodným restrikčním enzymem a pufrem (viz dodatek 6). Naštěpené fragmenty
se rozdělí elektroforézou v agarózovém gelu a pozoruje se charakteristický vzor restrikčních
fragmentů pod UV světlem po obarvení ethidiumbromidem a porovnává se s neštěpenou
a štěpenou pozitivní kontrolou.
Interpretace výsledků testu PCR
Test PCR je negativní, jestliže amplikon PCR specifický pro R. solanacearum
očekávané velikosti není u zkoumaného vzorku zjištěn, ale je zjištěn u všech pozitivních
kontrolních vzorků (v případě vícenásobné PCR s interními kontrolními primery specifickými
pro rostlinu: druhý produkt PCR očekávané velikosti musí být amplifikován se zkoumaným
vzorkem). Test PCR je pozitivní, jestliže je zjištěn amplikon PCR specifický pro
R. solanacearum očekávané velikosti a (případně) vzoru, za předpokladu, že není amplifikován
u žádného vzorku negativní kontroly. Spolehlivé potvrzení pozitivního výsledku lze
také získat opakováním testu s druhou sadou primerů PCR dodatek 6).
Poznámka:
Lze mít podezření na inhibici PCR, jestliže je získán očekávaný
amplikon ze vzorku pozitivní kontroly obsahujícího R. solanacearum ve vodě, zatímco
ze vzorku pozitivní kontroly R. solanacearum v bramborovém extraktu byly získány
negativní výsledky. V multiplexních protokolech PCR s interními kontrolami PCR nasvědčuje
inhibici reakce, jestliže není získán žádný ze dvou amplikonů.
V případě, že očekávaný amplikon je získán z jednoho nebo více vzorků negativních
kontrol, je podezření na kontaminaci.
6.1.7. Test FISH
Princip
Když se jako první screeningový test použije FISH test a je pozitivní,
musí být jako druhý povinný screeningový test provedena izolace nebo IF test. Když
se FISH test provede jako druhý vyšetřovací test a je pozitivní, je k dokončení diagnózy
nutné další testování podle postupového diagramu.
Poznámka:
Používají se schválené oligosondy specifické pro R. solanacearum
(dodatek 7). Úvodní testování touto metodou by mělo umožnit reprodukovatelné zjištění
alespoň 10
3
-104
buněk R. solanacearum na ml přidané do extraktů ze vzorku, které
byly předtím testovány s negativním výsledkem. Následující postup by měl být pokud
možno proveden s čerstvě připravenými extrakty, ale je možné jej úspěšně provést
s extraktem, který byl uchován v glycerolu při teplotě - 16 až - 24 °C nebo - 68
až - 86 °C.Jako negativní kontrola se použije alikvotní část extraktu ze vzorku, který
byl předtím testován na R. solanacearum s negativním výsledkem.
Jako pozitivní kontrola se připraví suspenze obsahující 10
5
až 106
buněk
na 1 ml R. solanacearum biovar 2 (např. kmen NCPPB 4156 = PD 2762 = CFBP 3857, viz
dodatek 3) v 0,01M fosfátovém pufru (PB) ze 3-5 denní kultury. Připraví se samostatná
sklíčka s pozitivními kontrolními vzorky homologického kmenu nebo jiného referenčního
kmene R. solanacearum, suspendovaného v bramborovém extraktu, jak je uvedeno v dodatku
3 B. Použití eubakteriálních oligosond značených FITC poskytuje kontrolu procesu
hybridizace, protože zbarví všechny eubakterie přítomné ve vzorku.Standardizovaný pozitivní a negativní kontrolní materiál, který se používá
v tomto testu, je uveden v dodatku 3 bodu A. Test kontrolního materiálu se provede
stejným způsobem jako u vzorku(ů).
6.1.7.1. Fixace bramborového extraktu
Následující protokol vychází z Wullingse et al. (1998).
1 Připraví se fixační roztok (viz dodatek 7).
2. Napipetuje se 100 µl každého vzorkového extraktu do Eppendorfovy mikrozkumavky
a odstřeďuje se po dobu 7 minut na 7 000 g.
3. Odstraní se supernatant a rozpustí se peleta ve 200 µl fixačního roztoku
připraveného max. 24 hodin předem. Protřepe se a inkubuje 1 hodinu v chladícím zařízení.
4. Odstřeďuje se 7 minut při 7 000 g, odstraní se supernatant a resuspenduje
se peleta v 75 µl 0,01M PB (viz dodatek 7).
5. Kápne se 16 µl fixované suspenze na čisté 10 okénkové sklíčko, jak ukazuje
obrázek 7.1, přičemž se použijí 2 různé vzorky na jedno sklíčko, a to neředěný a
zředěný 1:100 s použitím 10 µl (v 0,01 M PB). Zbývající roztok vzorku (49 µl) může
být uložen při teplotě - 20 °C po přidání 1 objemového množství 96 % etanolu. V případě,
že je třeba FISH metodu opakovat, odstraní se etanol odstředěním a přidá se stejné
množství 0,01 PB (zamíchá se protřepáním).

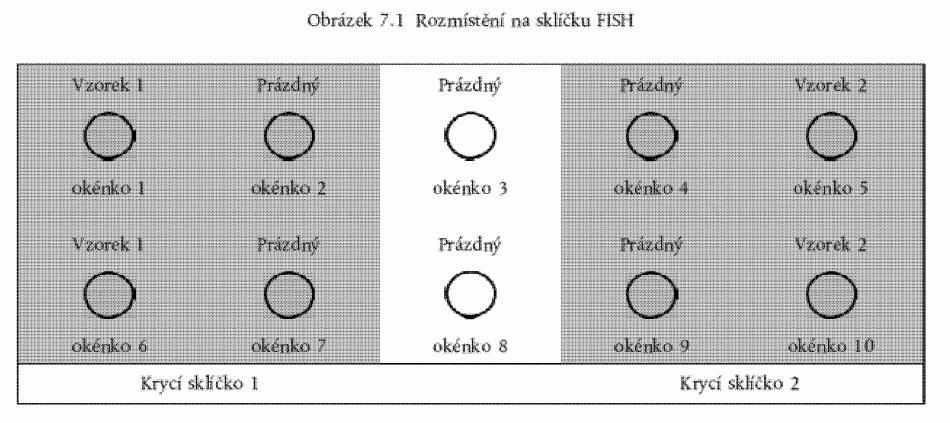
6. Sklíčka se nechají uschnout na vzduchu (nebo v sušičce sklíček při teplotě
37 °C) a fixují se nad plamenem.
V této fázi je možné postup přerušit a pokračovat v hybridizaci další den.
Sklíčka by měla být skladována chráněna před prachem a v suchu při pokojové teplotě.
6.1.7.2. Hybridizace
1. Dehydratují se buňky v postupné etanolové řadě 50 %, 80 % a 96 %, pokaždé
po dobu 1 minuty. Osuší se vzduchem v držáku sklíček.
2. Připraví se vlhká inkubační komora přikrytím dna vzduchotěsného boxu tkaninou
nebo filtračním papírem nasáklým hybridizační směsí (1x hybmix, dodatek 7). Před
inkubací se ohřeje box v hybridizační peci při teplotě 45 °C po dobu nejméně 10 minut.
3. Použije se 10 µl hybridizačního roztoku (dodatek 7) na 8 okének (okénka
1, 2, 4, 5, 6, 7, 9 a 10; viz obr.7.1) na každém sklíčku, přičemž dvě středová okénka
(3 a 8)se nechají prázdná.
4. Přiloží se krycí sklíčka (24 x 24 mm) na první a poslední 4 okénka, a
to tak, aby pod ně nevnikl vzduch. Sklíčka se umístí do předem zahřáté vlhké komory
a nechá se proběhnout proces hybridizace po dobu 5 hodin v troubě při teplotě 45
°C v temnu.
5. Připraví se 3 kádinky obsahující 1 l vody s molekulární kvalitou (Milli
Q), 1 l 1x hybmix (334 ml 3x hybmix a 666 ml vody s molekulární kvalitou) a 1 l 1/8x
hybmixu (42 ml 3x hybmix a 958 ml vody s molekulární kvalitou). Nechají se inkubovat
ve vodní lázni o teplotě 45 °C.
6. Sejmou se krycí sklíčka a podložní sklíčka se umístí do držáku sklíček.
7. Spláchne se nadbytek vzorku inkubací po dobu 15 minut v kádince s 1 x
hybmixem při teplotě 45 st. C.
8. Držák sklíček se přemístí do promývacího roztoku 1/8 hybmix a nechá se
inkubovat dalších 15 minut.
9. Sklíčka se krátce ponoří do UPW (např. Milli Q water) a položí na filtrační
papír. Odstraní se nadbytečná vlhkost lehkým zakrytím povrchu filtračním papírem.
Napipetuje se 5-10 µl krycího roztoku (např. Vectashield, Vecta Laboratories, CA,
USA nebo podobný) do každého okénka a celé sklíčko se zakryje velkým krycím sklíčkem
(24 x 60 mm).
6.1.7.3. Hodnocení FISH testu
1. Sklíčka se ihned prohlížejí pod mikroskopem vhodným pro epifluorescenční
mikroskopii se zvětšením 630 x nebo 1 000 x pod olejovou imerzí. S filtrem vhodným
pro fluorescein isothiokyanat (FITC) jsou eubakteriální buňky(včetně většiny gramnegativních
buněk) ve vzorku zbarveny fluorescenčně zeleně. Použitím filtru pro tetramethylrhodamin-5-
isothiokyanat se buňky R. solanacearum obarvené Cy3 jeví fluorescenčně červené. Porovnává
se buněčná morfologie s morfologií pozitivních kontrolních vzorků. Buňky musí být
jasně fluoreskující a zcela zbarveny. Test FISH (odst. 6.1.7) musí být zopakován,
pokud je zbarvení odchylné. Prohlížejí se okénka napříč dvěma průměry v pravých úhlech
a kolem obvodu. U vzorků, kde nejsou pozorovány žádné nebo málo buněk, se pozoruje
nejméně 40 polí mikroskopu.
2. Hledají se jasně fluoreskující buňky s morfologií charakteristickou pro
R. solanacearum v okénkách testovacích sklíček. Intenzita fluorescence musí odpovídat
nebo být lepší než u pozitivního kontrolního kmene. Buňky, které nejsou zcela zbarveny
nebo vykazují slabou fluorescenci, se neberou v úvahu.
3. Při podezření na jakoukoli kontaminaci musí být test opakován. To může
nastat v případě, kdy všechna sklíčka ve várce ukazují pozitivní buňky z důvodu kontaminace
pufru nebo jestliže jsou pozitivní buňky zjištěny (vně okénka sklíčka) na krytu sklíčka.
4. Specifičnost testu FISH s sebou nese několik problémů. Je pravděpodobné,
že u pletiv z bramborových hlíz a pelet ze stonkových partií bramboru dojde k výskytu
populací fluorescenčních buněk na pozadí s atypickou morfologií a ke křížové reakci
se saprofytickými bakteriemi velikostí a morfologií podobnými R. solanacearum, i
když mnohem méně častěji než u testu IF.
5. V úvahu se berou pouze fluoreskující buňky s typickou velikostí a morfologií.
6. Interpretace výsledků testu FISH
i) Výsledky FISH testu jsou platné, pokud jsou při použití FITC filtru
jasně zeleně fluoreskující buňky s velikostí a morfologií typickou pro R. solanacearum
a při použití rhodaminového filtru jasně červeně fluoreskující buňky pozorovány ve
všech pozitivních kontrolách a nejsou pozorovány v žádných negativních kontrolách.
Pokud jsou přítomné jasně fluoreskující buňky s typickou morfologií, odhadne se průměrný
počet typických buněk v 1 mikroskopovém poli a vypočítá počet typických buněk v 1
ml resuspendované pelety (dodatek 4). Vzorky, které obsahují alespoň 5 x 10
3
typických
buněk na 1 ml resuspendované pelety, se považují za pravděpodobně infikované. Nutné
je další testování. Vzorky, které obsahují méně než 5 x 103
typických buněk na 1
ml resuspendované pelety, se považují za negativní.ii) Výsledek FISH testu je negativní, pokud při použití rhodaminového filtru
nejsou pozorovány jasně červeně fluoreskující buňky s velikostí a morfologií typickou
pro R. solanacearum, jestliže jsou tyto typické jasně červeně fluoreskující buňky
při použití rhodaminového filtru pozorovány v pozitivních kontrolách.
6.1.8. Testy ELISA
Princip
Testy ELISA lze použít pouze jako volitelný test kromě testů IF,
PCR nebo FISH pro jeho relativně nízkou citlivost. Při použití DAS ELISA je obohacení
a použití monoklonálních protilátek povinné. Obohacení vzorků před použitím testu
ELISA může zvýšit citlivost testu, ale může se rovněž setkat s nezdarem kvůli konkurenci
jiných organismů ve vzorku.
Poznámka:
Použije se validovaný zdroj protilátek R. solanacearum. Doporučuje
se určení titru pro každou novou šarži protilátek. Titr je definován jako nejvyšší
ředění, při kterém dojde k optimální reakci při testování suspense obsahující 10
5
až 106
buněk na 1 ml homologického kmene R. solanacearum a použití vhodných druhotných
konjugátů protilátek podle doporučení výrobce. Při testování by měly být použity
protilátky v pracovním ředění, které je blízké nebo stejné jako u titru komerční
formulace.Určí se titr protilátek pro suspenzi 10
5
až 106
buněk na 1 ml homologického
kmene R. solanacearum.Vzorek, který byl předtím otestován jako R. solanacearum negativní a suspenze
bakterií bez vzájemného působení v solném roztoku fosfátového pufru (PBS) se použijí
jako vzorky negativní kontroly.
Jako pozitivní kontrolní vzorky se použijí alikvótní podíly vzorkových extraktů,
které byly předtím otestovány jako negativní, s příměsí 10
3
až 104
buněk na 1 ml
biovaru 2 R. solanacearum (např. kmen NCPPB 4156 = PD 2762 = CFBP 3857, viz dodatek
2 A a B). Pro srovnání výsledků na každé destičce se použije standardní suspenze
105
až 106
buněk na 1 ml v PBS R. solanacearum. Pozitivní kontrolní vzorky musí být
na mikrotitrové destičce dobře odděleny od vzorků k testování. Standardizované pozitivní
a negativní materiály, které se používají pro tento test, jsou uvedeny v dodatku
3 bodu A. Test kontrolního materiálu se provede týmž způsobem jako test vzorku(ů).Validované jsou dva protokoly ELISA.
a) Nepřímý test ELISA (Robinson Smith et al., 1995)
1) Použije se 100-200
µl vzorkového extraktu. (Zahřátí na 100 °C na 4 minuty ve vodní lázni nebo topném
boxu může v některých případech redukovat vznik nespecifických výsledků).
2) Přidá
se stejné množství dvojnásobně silného uhličitanového krycího pufru (dodatek 4) a
směs se promíchá.
3) Nakape se 100 µl do každé jamky mikrotitrační destičky (např.
Nunc-Polysorp nebo rovnocenné) a nechá se inkubovat 1 hodinu při teplotě 37 °C nebo
14 - 18 h při teplotě 4 °C.
4) Extrakty se vylijí z jamek. Jamky se třikrát vymyji
pomocí PBS-Tween (dodatek 4) a poslední vymývací roztok se nechá v jamce nejméně
5 minut.
5) Připraví se vhodné ředění protilátek proti R. solanacearum v blokačním
pufru (dodatek 4). U validovaných komerčních protilátek se použijí doporučená ředění
(obvykle dvojnásobné koncentrace, než je titr).
6) Přidá se 100 µl do každé jamky
a nechá se inkubovat 1 hodinu při teplotě 37 °C.
7) Roztok protilátek se vylije z
jamek a jamky se vymyjí jako předtím (bod 4.).
8) Připraví se vhodné ředění konjugátu
alkalické fosfatázy v blokačním pufru. Přidá se 100 µl do každé jamky a nechá se
inkubovat 1 hodinu při teplotě 37 °C.
9) Konjugát se vylije z jamek a jamky se vymyjí
jako předtím (bod 4).
10) Přidá se 100 µl substrátového alkalického roztoku fosfatázy
(dodatek 4) do každé jamky. Nechá se inkubovat v temnu při pokojové teplotě a odečítá
se absorbance při 405 nm v pravidelných 90minutových intervalech.
b) DAS-ELISA
1)
Připraví se vhodné ředění polyklonálního imunoglobulinu v uhličitanovém pufru pH
9.6 (dodatek 4). Přidá se 200 µl do každé jamky. Nechá se inkubovat při teplotě 37
°C 4 až 5 hodin nebo 4 ° C po dobu 16 hodin.
2) Jamky se třikrát dobře vypláchnou
použitím PBS-Tween (dodatek 4).Přidá se 190 µl extraktu ze vzorku do nejméně dvou
jamek. Přidají se rovněž pozitivní a negativní kontrolní vzorky do dvou jamek na
každé destičce. Nechá se inkubovat 16 hodin při teplotě 4 °C.
3) Jamky se třikrát
dobře vypláchnou použitím PBS-Tween (dodatek 4).
4) Připraví se vhodné ředění specifických
monoklonálních protilátek R. solanacearum v PBS (dodatek 4) s obsahem 0,5 % hovězího
sérového albuminu (BSA), přidá se 190 µl do každé jamky a nechá stát 2 hodiny při
teplotě 37 °C.
5) Jamky se třikrát dobře vypláchnou použitím PBS-Tween (dodatek 4).
6)
Připraví se ředění protimyšího imunoglobulinu konjugovaného alkalickou fosfatázou
v PBS. Přidá se 190 µl do každé jamky a nechá se inkubovat 2 hodiny při teplotě 37
°C.
7) Jamky se třikrát dobře vypláchnou použitím PBS-Tween (dodatek 4).
8) Připraví
se roztok 1 mg p-NPP/ml alkalické fosfatázy v substrátovém pufru (dodatek 4). Přidá
se 200 µl do každé jamky a inkubuje se v temnu při pokojové teplotě. Absorbance při
405 nm se odečítá v pravidelných intervalech 90 minut.
Interpretace výsledků testu ELISA
Test ELISA je negativní, jestliže průměrná hodnota optické hustoty (OH) z
jamek se stejnými vzorky je menší než dvojnásobek OH u negativního kontrolního vzorku,
pokud všechny hodnoty OH pozitivních kontrolních vzorků jsou větší než 1,0 (po 90
minutách inkubace se substrátem) a jsou větší než dvojnásobek OH získané z negativních
vzorkových extraktů. Test ELISA je pozitivní, jestliže průměrné hodnoty OH z jamek
se stejným vzorkem jsou větší než dvojnásobek OH v negativním extraktu testovaného
vzorku, pokud hodnoty OH ve všech negativních kontrolních vzorcích jsou menší než
dvojnásobek hodnot v pozitivních kontrolních vzorcích.
Negativní hodnoty ELISA v pozitivních kontrolních vzorcích ukazují, že test
nebyl proveden správně nebo že byl inhibován. Pozitivní hodnoty ELISA v negativních
kontrolních vzorcích ukazují, že došlo k vzájemné kontaminaci nebo nespecifickému
vázání protilátek.
6.1.9. Biotest
Poznámka.:
Předběžné testování touto metodou by mělo umožnit reprodukovatelnou
detekci 10
3
až 104
jednotek tvořících kolonie R. solanacearum na 1 ml přidaný do
extraktu ze vzorku, který byl předtím testován s negativním výsledkem (přípravu viz
v dodatku 3).Nejvyšší citlivost zjištění lze očekávat při použití čerstvě připraveného
extraktu ze vzorku a v optimálních růstových podmínkách. Metodu lze však také úspěšně
použít na extrakty, které byly uchovány v glycerolu v teplotě - 68 až - 86 °C. Následující
protokol vychází z Janseho (1988):
6.1.9.1. Použije se 10 testovacích rostlin citlivé odrůdy rajčete (např.
kultivaru Moneymaker nebo kultivaru s rovnocennou citlivostí podle testovací laboratoře)
ve fázi třech pravých listů u každého vzorku. Podrobnosti pěstování viz. v dodatku
8. Nebo se použije lilek (např. kultivar Black Beauty nebo kultivary s rovnocennou
citlivostí), ale jen rostliny ve fázi 2. až 3. listů až do plného rozvinu třetího
pravého listu. Příznaky u lilku jsou méně výrazné a vyvíjejí se pomaleji. Pokud možno,
doporučujeme proto použít sazenice rajčat.
6.1.9.2. Rozdělí se 100 µl extraktu ze vzorku mezi testovací rostliny.
1. Inokulace injekční stříkačkou
Inokulují se stonky rostlin hned nad děložními
listy pomocí stříkačky s podkožní jehlou (ne méně než 23G). Vzorek se rozdělí mezi
testovací rostliny.
2. Inokulace zářezem
Rostlina se přidrží mezi dvěma prsty a pipetou se kápne
přibližně 5-10 µl suspendované pelety na stonek mezi děložními listy a prvním listem.
Sterilním skalpelem se udělá příčný řez asi 1 cm dlouhý a hluboký přibližně
2/3 tloušťky stonku, přičemž řez se začne v místě kapky resuspendované pelety.
Řez se utěsní sterilní vazelínou z injekční stříkačky.
6.1.9.3. Stejnou technikou se nainokulujte 5 sazenic vodní suspenzí 10
5
až 106
buněk na 1 ml, připravenou ze 48 hodinové kultury virulentního kmene biovaru
2 R. solanacearum jako pozitivní kontrolní vzorek, a peletovým pufrem (dodatek 4)
jako negativní kontrolní vzorek. Oddělí se pozitivní a negativní kontrolní rostliny
od ostatních rostlin, aby se zabránilo vzájemné kontaminaci.6.1.9.4. Testovací rostliny se nechají růst v karanténním zařízení 4 týdny
při teplotě 25 -30 °C a vysoké relativní vlhkosti s přiměřeným zavlažováním, aby
se zabránilo jak přemokření, tak vadnutí z důvodu nedostatku vody. Aby se zabránilo
kontaminaci, musí být drženy pozitivní a negativní kontrolní rostliny ve zcela oddělených
místech skleníku nebo vegetační komory nebo jinak přísně izolovány. Musejí-li být
rostliny z různých vzorků po dobu inkubace drženy blízko sebe, oddělí se vhodnými
zástěnami. Při hnojení, zalévání, prohlížení a všech ostatních činnostech se věnuje
zvláštní pozornost tomu, aby nedošlo k vzájemné kontaminaci. Nutné je udržovat skleník
nebo vegetační prostor bez hmyzích škůdců, protože by mohli přenést bakterie mezi
vzorky.
Hledají se příznaky vadnutí, epinastie, chloróz anebo krnění rostlin.
6.1.9.5. Z infikovaných rostlin se provede izolace (odst. 2.3) a identifikují
se podezřelé čisté kultury (bod 6.2.).
6.1.9.6. Jestliže během 3 týdnů nejsou pozorovány žádné příznaky, provede
se test IF/PCR/izolace na složeném vzorku ze segmentů stonků dlouhých 1 cm z každé
testovací rostliny, odebraných nad místem inokulace. Je-li test pozitivní, provedou
se zřeďovací roztěry (bod 4.1).
6.1.9.7. Identifikují se všechny čisté kultury s podezřením na R. solanacearum
(bod 6.2.).
Interpretace výsledků biotestu
Platné výsledky biotestu jsou získány, jestliže pozitivní kontrolní testovací
rostliny vykazují typické příznaky, bakterie mohou být z těchto rostlin znovu izolovány
a negativní kontrolní rostliny nevykazují žádné příznaky. Biotest je negativní, jestliže
rostliny nejsou infikovány bakteriemi R. solanacearum, pokud byl organismus zjištěn
u pozitivních kontrolních testovacích rostlin. Biotest je pozitivní, jestliže testovací
rostliny jsou infikovány bakteriemi R. solanacearum.
6.2. Identifikační testy
Identifikují se čisté kultury izolovaného podezřelého organismu R. solanacearum
použitím nejméně dvou následujících testů založených na různých biologických principech.
Zahrnou se případně známé referenční kmeny pro každý provedený test (viz dodatek
3).
6.2.1. Výživové a enzymatické identifikační testy Určují se následující
fenotypové vlastnosti, které jsou vesměs přítomny nebo naopak chybí u R. solanacearum,
metodami Lelliott a Stead (1987), Klement et al. (1990), Schaad (2001).
Text Očekávaný výsledek Produkce fluorescenčního pigmentu - Inkluze PHB + Oxidačně fermentační test O+/F- Katalázová aktivita + Oxidázový test podle Kovacse + Redukce dusičnanů + Využití citrátů + Růst při 40 st. C - Růst v 1% NaCl + Růst v 2% NaCl - Arginin-dihydrolázová aktivita - Ztekucení želatiny - Hydrolýza škrobu - Hydrolýza eskulinu - Produkce levanu -
Média a metody viz. Lelliot &Stead (1987).
6.2.2. Test IF
6.2.2.1. Připraví se suspenze přibližně 106 buněk na 1 ml v pufru IF (dodatek
4).
6.2.2.2. Připraví se řada dvojnásobných zředění vhodného antiséra.
6.2.2.3. Použije se IF metoda (6.1.5.).
6.2.2.4. Test IF je pozitivní, jestliže titr IF kultury odpovídá titru pozitivní
kontroly.
6.2.3. Test ELISA
Poznámka:
Při provádění pouze 2 identifikačních testů se nepoužívá kromě
této metody jiný sérologický test.
6.2.3.1. Připraví se suspenze přibližně 10
8
buněk na 1 ml v 1X PBS (dodatek
4). 6.2.3.2. Provede se test ELISA se specifickou monoklonální protilátkou k
R. solanacearum.
6.2.3.3. Test ELISA je pozitivní, jestliže hodnota jeho výsledku získaná
z této kultury je nejméně poloviční v porovnání s hodnotou z pozitivní kontroly.
6.2.4. Testy PCR
6.2.4.1. Připraví se suspenze přibližně 106 buněk na 1 ml sterilní vody
(UPW = ultra pure water).
6.2.4.2. Ohřeje se 100 µl buněčné suspense v uzavřených zkumavkách v ohřívacím
bloku nebo vařící vodní lázni při 100 °C 4 minuty. Vzorky mohou být uchovány při
teplotě - 16 až - 24 °C do dalšího použití.
6.2.4.3. Použijí se příslušné postupy PCR k amplifikaci amplikonů specifických
pro R. solanacearum (např. Seal et al. (1993); Pastrik a Maiss (2000); Pastrik et
al. (2002); Boudazin et al. (1999); Opina et al. (1997), Weller et al. (1999).
6.2.4.4. Identifikace organismu R. solanacearum je pozitivní, jestliže amplikony
PCR mají stejnou velikost a mají stejnou mnohotvárnost délky fragmentů jako pozitivní
kontrolní kmen.
6.2.5. Test FISH
6.2.5.1. Připraví se suspenze přibližně 10
6
buněk na 1 ml v UPW. 6.2.5.2. Použije se metoda FISH (6.1.7) s nejméně 2 oligosondami specifickými
pro R. solanacearum (dodatek 7).
6.2.5.3. Test FISH je pozitivní, jestliže se u kultury a pozitivní kontroly
dosáhne stejných reakcí.
6.2.6. Analýza mastných kyselin (FAP)
6.2.6.1. Pěstuje se kultura na tryptikázo- sójovém agaru (Oxoid) 48 hodin
při teplotě 28 °C.
6.2.6.2. Použijte se metoda FAP (Janse, 1991; Stead, 1992).
6.2.6.3. Test FAP je pozitivní, jestliže je profil podezřelé kultury stejný
jako profil pozitivní kontroly. Výskyt charakteristických mastných kyselin: 14:0
3OH, 16:0 2OH, 16:1 2OH a 18:1 2OH a absence 16:0 3 OH poukazuje na Ralstonia sp.
6.2.7. Metody charakteristiky kmenů
Pro každý nový případ izolace R. solanacearum
se doporučuje charakteristika kmenu použitím jedné z následujících metod. U každého
provedeného testu se zahrnou případně známé referenční kmeny (viz. dodatek 3).
6.2.7.1. Stanovení biovaru
R. solanacearum tvoří biovary lišící se schopností využití a/nebo oxidace
tří disacharidů a tří hexosových alkoholů (Hayward, 1964 a Hayward et al., 1990).
Živná půda pro test biovaru je popsána v dodatku 2. Test lze úspěšně provést očkováním
do živné půdy s čistou kulturou R. solanacearum a inkubací při teplotě 28 °C. Je-li
živná půda rozdělena na sterilní destičky s 96 jamkami (200 µl na 1 jamku), zbarvení
se mění během 72 hodin od olivově zelené po žlutou a znamená pozitivní výsledek testu.

6.2.7.2. Genomická variabilita
Molekulární identifikace kmenů komplexu R. solanacearum lze dosáhnout několikerými
technikami, mezi něž patří:
1. Analýza délkového polymorfismu restrikčních fragmentů RFLP (Cook et al.,
1989).
2. Repetitivní sekvenční PCR s využitím primerů REP, BOX a ERIC (Louws et
al., 1995; Smith et al., 1995).
3. Analýza délkového polymorfismu amplifikovaných fragmentů AFLP (Van der
Wolf et al., 1998).
6.2.7.3. Metody PCR
Specifické primery PCR (Pastrik et al, 2002; viz dodatek 6) lze použít k
diferenciaci kmenů patřících do skupiny 1 (biovary 3, 4 a 5) a skupiny 2 (biovary
1, 2A a 2T) R. solanacearum, jak bylo původně definováno analýzou RFLP (Cook et al.,
1989) a sekvenováním 16S rDNA (Taghavi et al., 1996).
6.3. Konfirmační test (test patogenity)
Jako závěrečné potvrzení diagnózy R. solanacearum a k potvrzení virulence
kultur identifikovaných jako R. solanacearum musí být proveden test patogenity.
1) Připraví se inokulum o hustotě přibližně 106 buněk na 1 ml ze 24 až 48
hodinových kultur k testování a příslušný kmen pozitivní kontroly R. solanacearum
(např. NCPPB 4156 = PD 2762 = CFBP 3857; viz dodatek 3).
2) Inokuluje se 5 až 10 náchylných sazenic rajčete nebo lilku ve fázi 3 pravých
listů (viz odst. 6.1.9).
3) Sazenice se nechají inkubovat až 2 týdny při teplotě 25-28 °C a vysoké
relativní vzdušné vlhkosti s přiměřeným zaléváním, bez vystavení zamokření nebo vyprahlosti
půdy. U čistých kultur by typické vadnutí mělo nastat během 14 dnů. Neobjeví-li se
po této době příznaky, kultura nemůže být považována jako patogenní forma R. solanacearum.
4) Hledají se příznaky vadnutí a/nebo epinastie, chlorózy nebo krnění.
5) Provede se izolace z rostlin vykazujících příznaky odebráním segmentu
ze stonku asi 2 cm nad místem inokulace. Segment se rozmělní a suspenduje v malém
množství sterilní destilované vody nebo 50 mM fosfátového pufru (dodatek 4). Izolace
ze suspense se provede zřeďovacím roztěrem pokud možno na selektivní živné půdě (dodatek
2). Po inkubaci 48 až 72 hodin při teplotě 28 °C se vyhodnotí výskyt typických kolonií
pro R. solanacearum.
Dodatek 1
Laboratoře podílející se na optimalizaci a validaci protokolů
------------------------------------------------ ---------------------------------- Laboratoř (1) Místo Země ------------------------------------------------ ---------------------------------- Agentur für Gesundheit und Ernährungssicherheit Vídeň a Linec Rakousko Departement Gewasbescherming Merelbeke Belgie Plantedirektoratet Lyngby Dánsko Central Science Laboratory York Anglie Scottish Agricultural Science Agency Edinburgh Skotsko Laboratoire National de la Protection Végétaux, Unité de Bactériologie Angers Francie Laboratoire National de la Protection Végétaux, Station de Quarantaine de la Pomme de Terre Le Rheu Francie Biologische Bundesanstalt Kleinmachnow Německo Pflanzenschutzamt Hannover Hannover Německo State Laboratory Dublin Irsko Dipartimento di Scienze e Tecnologie Agroambientali Boloň Itálie Regione Veneto Unita Periferica per i Servizi Fitosanitari Verona Itálie Nederlandse Algemene Keuringsdienst Emmeloord Nizozemsko Plantenziektenkundige Dienst Wageningen Nizozemsko Direcçao-General de Protecçao das Culturas Lisabon Portugalsko Centro de Diagnóstico de Aldearrubia Salamanca Španělsko Instituto Valenciano de Investigaciones Agrarias Valencie Španělsko Swedish University of Agricultural Sciences Uppsala Švédsko ------------------------------------------------ ---------------------------------- (1) Kontakty: viz internetová stránka http://forum.europa.eu.int/Public/irc/sanco/Home/main
Dodatek 2
Média pro izolaci a kultivaci organismu R. solanacearum
a) Živné půdy pro izolaci a kultivaci
Živný agar (NA)
Živný agar (Difco) 23,0 g
Destilovaná voda 1,00 l
Rozpustí se složky a sterilují tlakem při teplotě 121 °C po dobu 15 minut.
Kvasnično-pepton-glukózový agar (YPGA)
Kvasnicový extrakt (Difco) 5,0 g
Bacto-Pepton (Difco) 5,0 g
D(+) Glukóza (monohydrát) 10,0 g
Bacto-Agar (Difco) 15,0 g
Destilovaná voda 1,00 l
Rozpustí se složky a sterilují tlakem při teplotě 121 °C po dobu 15 minut.
Sacharózopeptonový agar (SPA)
Sacharóza 20,0 g
Bacto-Peptone (Difco) 5,0 g
K2 HPO4 0,5 g
MgSO4 . 7H2O 0,25 g
Bacto-Agar (Difco) 15,0 g
Destilovaná voda 1,00 l
pH 7,2-7,4
Rozpustí se složky a sterilují v autoklávu při teplotě 121 °C po dobu 15 minut.
Kelmanovo tetrazoliové médium
kasein hydrolyzát (Difco) 1,0 g
Bacto-Pepton (Difco) 10,0 g
Dextróza 5,0 g
Bacto-Agar (Difco) 15,0 g
Destilovaná voda 1,00 l
Rozpustí se složky a sterilují v autoklávu při teplotě 121 °C po dobu 15 minut.
Ochladí se na 50 °C a přidá se filtrem sterilizovaný
roztok 2,3,5-trifenyl-tetrazoliumchloridu (Sigma), až do
dosažení konečné koncentrace 50 mg na litr.
b) Validované selektivní živné půdy
Živná půda SMSA (Englebrecht, 1994 ve znění Elphinstone et al., 1996)
Základní živná půda
Casamino kyseliny (Difco) 1,0 g
Bacto-Pepton (Difco) 10,0 g
Glycerol 5,0 ml
Bacto-Agar (Difco) (viz pozn. 2) 15,0 g
Destilovaná voda 1,00 l
Rozpustí se složky a sterilují v autoklávu při teplotě 121 °C po dobu 15 minut.
Ochladí se na 50 °C a přidá filtrem sterilizovaný
vodný roztok následujících složek, aby se získala stanovená
konečná koncentrace:
Krystalová violeť (Sigma) 5 mg/l
Polymixin-B-sulfát (Sigma P1004) 600 000 U (přibližně 100 mg)/l
Bacitracin (Sigma B-0125) 1 250 U (přibližně 25 mg)/l
Chloramfenikol (Sigma C-3175) 5 mg/l
Penicilín-G (Sigma P-3032) 825 U (přibližně 0,5 mg)/l
2,3,5-trifenyltetrazolium chlorid (Sigma) 50
mg/l
Poznámky:
1. Použití jiných než výše uvedených reagencí může ovlivnit růst
R. solanacearum.
2. Místo Bacto-Agar (Difco) může být použit Oxoid Agar #1. V takovém
případě bude růst R. solanacearum pomalejší, ale může to také omezit růst konkurenčních
saprofytů. Typické kolonie R. solanacearum se mohou tvořit o 1-2 dny déle a červené
zbarvení může být světlejší a rozptýlenější než při použití Bacto-Agaru.
3. Zvýšení
koncentrace bacitracinu na 2 500 U/l může omezit populace konkurenčních bakterií
bez ovlivnění růstu Ralstonia solanacearum.
Roztoky antibiotik se uchovávají a skladují při teplotě 4 °C v temnu a musí
se spotřebovat do 1 měsíce.
Misky s médiem by měly být před použitím zbaveny povrchové kondenzace.
Nutno zabránit přílišnému vysušení média. Každá nově připravená šarže živné
půdy by měla být podrobena kontrole kvality živné půdy roztěrem suspense referenční
kultury R. solanacearum na médium (viz dodatek 3) a pozorováním, zda se po 2 až 5
dnech při teplotě 28 °C objeví typické kolonie.
c) Validované obohacené živné půdy
Živná půda SMSA (Elphinstone et al., 1996) Provede se příprava jako pro selektivní
agarovou živnou půdu SMSA, ale vynechá se Bacto-Agar a 2,3,5- trifenyltetrazolium
chlorid.
Upravená živná půda Wilbrink (Caruso et al., 2002)
Sacharóza 10 g
Proteázový pepton 5 g
K2 HPO4 0,5 g
MgSO4 0,25 g
NaNO3 0,25 g
Destilovaná voda 1,00 l
Steriluje se tlakem při teplotě 121 °C po dobu 15 minut a ochladí se na 50 °C.
Přidá se antibiotický zásobní roztok jako pro živnou půdu SMSA.
Dodatek 3
A. Komerční standardizovaný kontrolní materiál
a) Izolované bakteriální kultury Následující izolované bakteriální kultury
se doporučují jako standardní referenční materiál buď k pozitivní kontrole (tabulka
1) nebo během optimalizace testů, aby se zabránilo vzájemnému působení (tabulka 2).
Všechny kmeny jsou komerčně dostupné a lze je získat z těchto sbírek:
1. Národní sbírka patogenních bakterií rostlin (NCPPB), Ústřední vědecká
laboratoř York, UK.
2. Sbírka kultur sekce ochrany rostlin (PD), Wageningen, Nizozemsko.
3. Francouzská sbírka patogenních bakterií rostlin (CFBP), Ústav fytobakteriologie
INRA, Angers, Francie.
Tabulka 1: Referenční panel izolovaných bakteriálních kultur SMT R. solanacearum
-------------------------------------------------------- --------------------------
Kód NCPPB SMT
# Jiné kódy Země původu Biovar
-------------------------------------------------------- --------------------------
NCPPB 4153 6 CFBP 4582, Pr 3020, EURS11 Egypt 2
NCPPB 4154 10 CFBP 4585, 550, EURS21 Turecko 2
NCPPB 3857 12 CFBP 4587, Pr 1140, EURS26 Anglie 2
NCPPB 1584 23 CFBP 4598, EURS49 Kypr 2
NCPPB 2505 24 CFBP 4599, EURS50 Švédsko 2
NCPPB 4155 26 CFBP 4601, 502, EURS55 Belgie 2
NCPPB 4156 (*) 71 (*) PD 2762, CFBP 3857 Nizozemsko 2
NCPPB 4157 66 LNPV 15.59 Francie 2
NCPPB 4158 39 CFBP 4608, Port 448, EURS80 Portugalsko 2
NCPPB 4160 69 IVIA-1632-2 Španělsko 2
NCPPB 4161 76 B3B Německo 2
NCPPB 325 41 CFBP 2047, KEL60-1, R842 USA 1
NCPPB 3967 42 CFBP 4610, R285, GONg7 Kostarika 1
NCPPB 4028 43 CFBP 4611, R303/571, CIP310,
SEQ205 Kolumbie 2
NCPPB 3985 44 CFBP 4612, R578, CIP312 Peru 2T
NCPPB 3989 45 CFBP 4613, R568, CIP226 Brazílie 2T
NCPPB 3996 46 CFBP 3928, R276/355, CIP72,
SEQ225 Peru 3
NCPPB 3997 47 CFBP 4614, R280/363, CIP49,
HAY0131 Austrálie 3
NCPPB 4029 48 CFBP 4615, R297/349, CIP121,
CMIb2861 Srí Lanka 4
NCPPB 4005 49 CFBP 4616, R470 Filipíny 4
NCPPB 4011 50 CFBP 4617, R288, HEmps2 Čína 5
-------------------------------------------------------- --------------------------
(*) Jako standardní referenční kmen se použije R. solanacearum
biovar 2 (rod 3).
Poznámka:
Spolehlivost výše uvedených kmenů může být zaručena pouze v případě,
že pocházejí z původních sbírek kultur.
Tabulka 2: Referenční panel SMT sérologicky nebo geneticky
příbuzných bakterií k optimalizaci detekčních testů
-------------------------------------- --------------------------------------
Kód NCPPB SMT
# Jiné kódy Identifikace
-------------------------------------- --------------------------------------
NCPPB 4162 51 CFBP 1954 Bacillus polymyxa (1)
NCPPB 4163 52 CFBP 1538 Pseudomonas marginalis
pv. marginalis (1)
NCPPB 4164 - CFBP 2227 Burkholderia cepacia (2)
NCPPB 4165 - CFBP 2459 Ralstonia pickettii (2)
NCPPB 4166 58 CFBP 3567,
CSL Pr1150 Ralstonia pickettii (1)
NCPPB 4167 60 CFBP 4618,
PD 2778 Ralstonia sp. (1)
NCPPB 1127 53 CFBP 3575 Burkholderia andropogonis (1)
NCPPB 353 54 CFBP 3572 Burkholderia caryophylli (1)
NCPPB 945 55 CFBP 3569 Burkholderia cepacia (1)
NCPPB 3708 56 CFBP 3574 Burkholderia glumae (1)
NCPPB 3590 57 CFBP 3573 Burkholderia plantarii (1)
NCPPB 3726 59 CFBP 3568 Banana Blood Disease Bacterium (1) (2) (3)
NCPPB 4168 61 CFBP 4619,
IPO S339 Enterobacter sp. (1)
NCPPB 4169 62 IPO 1695 Enterobacter sp. (1)
NCPPB 4170 63 CFBP 4621,
IPO S306 Ochrobactrum anthropi (1) (2)
NCPPB 4171 64 CFBP 4622,
IPO 1693 Curtobacterium sp. (1) (2)
NCPPB 4172 65 IPO 1696a Pseudomonas sp. (1)
NCPPB 4173 - PD 2318 Aureobacterium sp. (2)
NCPPB 4174 81 IVIA 1844.06 Flavobacterium sp. (1) (2)
-------------------------------------- --------------------------------------
(1) Potencionální křížově reagující kmen v sérologických testech (IF
a/nebo ELISA) s polyklonálním antisérem.
(2) Kmen, ze kterého lze v některých laboratořích amplifikovat produkt
PCR na podobnou velikost, jako je očekávaná velikost při použití specifických primerů
OLI-1 a Y- 2 (viz dodatek 6).
(3) Pravděpodobnost křížové reakce ve většině testů, ale výskyt znám
pouze u banánů v Indonésii.
b) Komerční standardizovaný kontrolní materiál
Následující standardní kontrolní materiál je možné získat ze sbírky kultur
NCPPB.
Mražené sušené pelety bramborového extraktu ze 200 zdravých hlíz bramboru
k negativní kontrole všech testů. Mražené sušené pelety bramborového extraktu ze
200 zdravých hlíz bramboru obsahující 10
3
až 104
a 104
až 106
buněk biovaru 2 R.
solanacearum (kmen NCPPB 4156 = PD 2762 = CFBP 3857) jako pozitivní kontroly serologických
a PCR testů. Protože životaschopnost buňky je ovlivněna lyofilizací, nejsou tyto
vhodné jako standardy v testech izolace a testech patogenity. Ve formaldehydu fixované
suspense biovaru 2 R. solanacearum (kmen NCPPB 4156 = PD 2762 = CFBP 3857) s 106
buňkami na 1 ml k pozitivní kontrole sérologických testů.B. Příprava pozitivních a negativních kontrol pro screeningové testy vodivých
pletiv PCR/IF a FISH
Kultivuje se 48hodinová kultura virulentního kmene R. solanacearum rasy 3/biovaru
2 (např. kmen NCPPB 4156 = PD 2762 = CFBP 3857) na SMSA živné půdě a suspenduje se
v 10 mM fosfátovém pufru, aby se získala buněčná hustota přibližně 2 × 108 buněk
tvořících kolonie na 1 ml. Obvykle se jí dosáhne jemně zakalenou suspenzí rovnající
se optické hustotě 0,15 při 600 nm.
Odebere se dřeň z pupkových konců 200 hlíz z produkce odrůdy s bílou slupkou,
o které se ví, že je prosta R. solanacearum. Zpracují se pupkové konce výše uvedeným
postupem a resuspenduje se peleta v 10 ml.
Připraví se 10 sterilních mikrozkumavek o objemu 1,5 ml s 900 µl resuspendované
pelety.
Přenese se 100 µl suspenze R. solanacearum do první mikrozkumavky a protřepe
se..
Provede se desetinásobné ředění v dalších pěti mikrozkumavkách.
Těchto šest kontaminovaných mikrozkumavek se použije jako kontrolní vzorky.
Čtyři nekontaminované mikrozkumavky se použijí jako kontrolní negativní vzorky. Mikrozkumavky
musí být řádně označeny.
Připraví se alikvotní části z 100 µl ve sterilních zkumavkách o objemu 1,5
ml, čímž se získá 9 kopií každého kontrolního vzorku. Uskladní se při teplotě - 16
až - 24 °C až do doby použití.
Přítomnost a množství R. solanacearum v kontrolních vzorcích by měly být
nejprve potvrzeny testem IF.
Pro PCR test se provede extrakce DNA z pozitivních a negativních kontrolních
vzorků pro každou sérii zkušebních vzorků.
Pro IF a FISH testy se provede test patogenity pozitivních a negativních
kontrolních vzorků pro každou sérii zkušebních vzorků.
Při kvantitativních rozborech IF, FISH a PCR musí být R. solanacearum zjištěna
v nejméně v 106 a 10 4 buněk/ml pozitivních kontrol a nesmí být zjištěn v žádné z
negativních kontrol.
Dodatek 4
Pufry pro testovací postupy
OBECNĚ: Neotevřené sterilizované pufry lze skladovat po dobu až jednoho roku.
1. Pufry pro extrakci
1.1 Extrakční pufr (50 mM fosfátový pufr, pH 7,0)
Tento pufr se používá k extrakci bakterie z rostlinných tkání homogenizací
nebo protřepáním.
Na2 HPO4 (bezvodý) 4,26 g
KH2PO4 2,72 g
Destilovaná voda 1,00 l
Rozpustí se složky, zkontroluje pH a sterilizuje v autoklávu při 121 °C po
dobu 15 min.
Užitečné mohou být následující složky:
Účel Množství (na litr)
Lubrolové vločky Protisrážlivý prostředek (*) 0,5 g
DC silikonový odpěňovač Odpěňovací činidlo (*) 1,0 ml
Tetrasodiumpyrofosfát Antioxidační činidlo 1,0 g
Polyvinylpyrrolidon-40 000 (PVP - 40) Vázání inhibitorů PCR 50 g
---------------------------------------
(*) pro použití při extrakci homogenizací
1.2. Peletový pufr (10 mM fosfátový pufr, pH 7,2)
Tento pufr se používá pro resuspenzi a ředění extraktů z výkrojků z pupkových
částí bramborových hlíz poté, co byly odstřeďováním koncentrovány do pelety.
Na2HPO4 .12H2O 2,7 g
NaH2PO4 . 2H2O 0,4 g
Destilovaná voda 1,00 l
Rozpustí se složky, zkontroluje pH a sterilizuje v autoklávu při 121 °C po
dobu 15 min.
2. Pufry pro IF test
Pufr pro IF (10 mM fosfátový pufr ve fyziologickém roztoku (PBS), pH 7,2)
Tento pufr se používá k ředění protilátek.
Na2HPO4 .12H2O 2,7 g
NaH2PO4 . 2H2O 0,4 g
NaCl 8,0 g
Destilovaná voda 1,00 l
Rozpustí se složky, zkontroluje pH a sterilizuje v autoklávu při 121 °C po
dobu 15 min.
IF-pufr-Tween
Tento pufr se používá k oplachování podložních sklíček. Přidá
se 0,1 % Tween 20 k pufru pro IF.
Fosfátový pufr v glycerolu, pH 7,6
Tento pufr se používá jako krycí roztok na okénka sklíček na IF testy k zvýšení
fluorescence.
Na2HPO4 .12H2O 3,2 g
NaH2PO4 . 2H2O 0.15 g
Glycerol 50 ml
Destilovaná voda 100 ml
Krycí roztoky jsou komerčně dostupné, např. Vectashield® (Vector Laboratories)
nebo Citifluor(R) (Leica).
3. Pufry pro účely nepřímého testu ELISA
3.1. Dvojnásobný uhličitanový potahovací pufr, pH 9,6.
Na2CO3 6,36 g
NaH CO3 11,72 g
Destilovaná voda 1,00 l
Rozpustí se složky, zkontroluje pH a steriluje autoklávováním při teplotě
121 °C po dobu 15 minut.
Pokud extrakt obsahuje vysoký podíl aromatických molekul, je možné jako antioxidant
přidat siřičitan sodný (0,2 %).
3.2. Desetinásobný roztok chloridu sodného pufrovaný fosfátem (10 x PBS),
pH 7,4
NaCl 80,0 g
KH2PO4 2,0 g
Na2HPO4 . 12H2O 29,0 g
KCl 2,0 g
Destilovaná voda 1,00 l
3.3. PBS-Tween
10 x PBS 100 ml
10 % Tween 20 5 ml
Destilovaná voda 895 ml
3.4. Blokační pufr (musí být čerstvě připraven).
10 x PBS 10,0 ml
Polyvinylpyrrolidon-44000
(PVP-44) 2,0 g
10 % Tween 20 0,5 ml
Sušené mléko 0,5 g
Destilovaná voda doplnit do 100 ml
3.5. Roztok pro substrát alkalické fosfatázy, pH 9,8
Diethanolamin 97 ml
Destilovaná voda 800 ml
Rozpustí se a koncentrovanou HCl se upraví pH na 9,8.
Doplní se do 1 l destilovanou vodou.
Přidá se 0,2 g MgCl2.
Rozpustí se dvě 5 mg tabletky fosfatázového substrátu (Sigma) v 15 ml roztoku.
4. Pufry pro účely testu DASI ELISA
4.1. Potahovací pufr, pH 9,6.
Na2CO3 1,59 g
NaH CO3 2,93 g
Destilovaná voda 1 000 ml
Rozpustí se složky a zkontroluje pH 9,6.
4.2. 10 x fosfátosolný pufr (PBS) pH 7,2- 7,4
NaCl 80,0 g
Na2HPO4 .12H2O 27 g
NaH2PO4 . 2H2O 4 g
Destilovaná voda 1 000 ml
4.3. PBS-Tween
10 x PBS 50 ml
10 % Tween 20 5 ml
Destilovaná voda 950 ml
4.4 Substrátový pufr, pH 9,8
Diethanolamin 100 ml Destilovaná voda 900 ml
Smíchá se a nastaví se hodnota pH 9,8 koncentrovanou HCl.
Dodatek 5
Stanovení koncentrace IF a FISH pozitivních buněk
1. Vypočítá se průměrný počet typických fluoreskujících buněk v jednom pozorovacím
poli (c).
2. Vypočítá se počet typických fluoreskujících buněk v okénku mikroskopického
sklíčka (C).
C = c x S/s,
kde S = plocha jednoho pole na sklíčku s více okénky a
s
= plocha pole objektivu.
s = píi
2
/4G2K 2,kde i = koeficient pole (v rozmezí 8 - 24
podle typu okuláru),
K = tubusový koeficient (1 nebo 1,25),
G = zvětšení objektivu
(100 x, 40 x atd.).
3. Vypočítá se počet charakteristických fluoreskujících buněk na 1 ml resuspendované
pelety (N).
N =C x 1 000/y x F,
kde y = objem resuspendované pelety v každém okénku
a
F = zřeďovací faktor resuspendované pelety
Dodatek 6
Validované protokoly a činidla pro PCR
Poznámka:
Úvodní testování by mělo umožnit reprodukovatelné zjištění nejméně
10
3
až 104
buněk R. solanacearum na 1 ml vzorkového extraktu. Úvodní testování by
také nemělo vykazovat žádné falešné pozitivní výsledky se skupinou vybraných kmenů
bakterií (viz dodatek 3).1. Protokol PCR Seal et al. (1993)
1.1. Oligonukleotidové primery
Přímý primer OLI-1: 5'-GGG GGT AGC TTG CTA CCT GCC-3'
Reverzní primer Y-2: 5'-CCC ACT GCT GCC TCC CGT AGG AGT-3'
Očekávaná velikost amplikonu templátové DNA R. solanacearum = 288 bp
1.2. Reakční směs PCR
Reagencie Množství v reakci Konečná koncentrace Sterilní UPW 17,65 µl 10x pufr PCR (1) (15 mM MgCl2 ) 2,5 µl 1x (1,5 mM MgCl2) Směs dNTP (20mM) 0,25 µl 0,2 mM Primer OLI-1 (20 µM) 1,25 µl 1µM Primer Y-2 (20 µM) 1,25 µl 1µM Taq polymerasa (5U/µl)(1) 0,1 µl 0,5 U Množství vzorku 2,0 µl Celkové množství 25 µl
(1) Metoda byla validována použitím Taq polymerázy Perkin Elmer (AmpliTaq)
a Gibco BRL.
1.3. Reakční podmínky PCR
Provede se následující cyklický proces:
1 cyklus: i) 2 minuty při 96 °C (denaturace templátové DNA)
35 cyklů: ii) 20 sekund při 94 °C (denaturace templátové DNA)
iii) 20 sekund při 68 °C (hybridizace primerů s templátovou DNA)
iv) 30 sekund při 72 °C (syntéza kopie)
1 cyklus: v) 10 minut při 72 °C (závěrečná syntéza)
vi) ponechá se při teplotě 4 °C.
Poznámka:
Tento program byl optimalizován pro použití termocykleru Perkin
Elmer 9600. Použití jiných přístrojů může vyžadovat úpravu jednotlivých kroků cyklu
ii), iii) a iv).
1.4. Restrikční enzymová analýza amplikonu.
Produkty PCR vzniklé amplifikací z R. solanacearum DNA vytvářejí polymorfismus
délky restrikčních fragmentů s enzymem Ava II po inkubaci při teplotě 37 °C.
2. Protokol PCR Pastrika a Maisse (2000)
2.1. Oligonukleotidové primery
Přímý primer Ps-1: 5'-agt cga acg gca gcg ggg g - 3'
Reverzní primer Ps-2: 5'-ggg gat ttc aca tcg gtc ttg ca -3'
Očekávaná velikost amplikonu templátové DNA R. solanacearum DNA = 553 bp.
2.2. Reakční směs PCR
Reagencie Množství v reakci Konečná koncentrace Sterilní UPW 16,025 µl 10x pufr PCR (1) 2,5 µl 1x (1,5 mM MgCl2) BSA (frakce V) (10 %) 0,25 µl 0,1 % Směs dNTP (20mM) 0,125 µl 0,1 mM Primer Ps-1 (10 µM) 0,5 µl 0,2 µM Primer Ps-2 (10 µM) 0,5 µl 0,2 µM Taq polymerasa (5U/µl)(1) 0,1 µl 0,5 U Množství vzorku 5,0 µl Celkové množství 25,0 µl
(1) Metoda byla validována použitím Taq polymerázy Perkin Elmer (AmpliTaq)
a Gibco BRL.
Poznámka:
Původně optimalizovaná pro termocykler MJ Research PTC 200 s polymerasou
Gibco Taq Polymerace. Perkin Elmer AmpliTaq a pufr mohou být rovněž použity ve stejných
koncentracích.
2.3 Reakční podmínky PCR
Provede se následující cyklický proces:
1 cyklus i) 5 minut při 95 °C (denaturace templátové DNA)
35 cyklů: ii) 30 sekund při 95 °C (denaturace templátové DNA)
iii) 30 sekund při 68 °C (hybridizace primerů s templátovou DNA)
iv) 45 sekund při 72 °C (syntéza kopie)
1 cyklus: v) 5 minut při 72 °C (syntéza extenze)
vi) ponechá se při teplotě 4 °C.
Poznámka:
Tento program byl optimalizován pro použití termocykleru MJ Research
PTC 200. Použití jiných přístrojů bude možná vyžadovat úpravu jednotlivých kroků
cyklu ii), iii) a iv).
2.4. Restrikční enzymová analýza amplikonu
Produkty PCR vzniklé amplifikací
z R. solanacearum DNA vytvářejí zřetelný polymorfismus délky restrikčních fragmentů
s enzymem Taq I po inkubační době 30 minut při teplotě 65 °C. Restrikční fragmenty
specifické pro R. solanacearum mají velikost 457 bp a 96 bp.
3. Protokol pro multiplexní PCR s interní kontrolou (Pastrik et al., 2002)
3.1. Oligonukleotidové primery
Přímý primer RS-1-F: 5'-ACT AAC GAA GCA GAG ATG CAT TA-3'
Reverzní primer RS-1-R: 5'-CCC AGT CAC GGC AGA GAC T-3'
Přímý primer NS-5-F: 5'-AAC TTA AAG GAA TTG ACG GAA G-3'
Reverzní primerNS-6-R: 5'-GCA TCA CAG ACC TGT TAT TGC CTC-3'
Očekávaná velikost amplikonu templátové DNA R. solanacearum = 718 bp (sada
primerů RS).
Očekávaná velikost amplikonu interní kontroly PCR 18S rRNA = 310 bp (sada
primerů NS).
3.2. Reakční směs PCR
Reagencie Množství v reakci Konečná koncentrace Sterilní UPW 12,625 µl 10x pufr PCR (1) (15 mM MgCl2) 2,5 µl 1x (1,5 mM MgCl2) BSA (frakce V) (10 %) 0,25 µl 0,1 % Směs dNTP (20mM) 0,125 µl 0,1 mM Primer RS-1-F (10 µM) 2,0 µl 0,8 µM Primer RS-1-R (10 µM) 2,0 µl 0,8 µM Primer NS-S-F (10 µM) (2) 0,15 µl 0,06 µM Primer NS-6-R (10 µM) (2) 0,15 µl 0,06 µM Taq polymeráza (5U/µl)(1) 0,2 µl 1,0 U Množství vzorku 5,0 µl Celkové množství 25,0 µl
-----
(1) Metoda byla validována použitím Taq polymerázy Perkin Elmer (AmpliTaq)
a Gibco BRL.
(2) Koncentrace primerů NS-5-F a NS-6-R byly optimalizovány pro výkrojky
pletiva pupkových částí hlíz bramboru pomocí homogenizační metody a purifikace DNA
podle Pastrika (2000) (viz 6.1.6.1.a)). Použití extrakční metody třepáním nebo jiných
metod izolace DNA může vyžadovat nové provedení optimalizace koncentrací reagencií.
3.3. Reakční podmínky PCR
Provede se následující cyklický proces:
1 cyklus: i) 5 minut při 95 °C (denaturace templátové DNA)
35 cyklů: ii) 30 sekund při 95 °C (denaturace templátové DNA)
iii) 30 sekund při 58 °C (hybridizace primerů s templátovou DNA)
iv) 45 sekund při 72 °C (syntéza kopie)
1 cyklus: v) 5 minut při 72 °C (konečná syntéza)
vi) nechá se při teplotě 4 °C.
Poznámka:
Tento program byl optimalizován pro použití termocykleru MJ Research
PTC 200. Použití jiných přístrojů může vyžadovat úpravu jednotlivých kroků cyklu
ii), iii) a iv).
3.4. Restrikční enzymová analýza amplikonu
Produkty PCR vzniklé amplifikací z DNA R. solanacearum vytvářejí zřetelný
polymorfismus délky restrikčních fragmentů s enzymem Bsm I nebo Isoschizomere (např.
Mva 1269 I) po inkubační době 30 minut při teplotě 65 °C.
4. Protokol PCR specifický pro biovar R. solanacearum (Pastrik et al, 2001)
4.1. Oligonukleotidové primery
Přímý primer Rs-1-F: 5'-ACT AAC GAA GCA GAG ATG CAT TA-3' Reverzní primer Rs-1-R: 5'-CCC AGT CAC GGC AGA GAC T-3' Reverzní primer Rs-3-R: 5'-TTC ACG GCA AGA TCG CTC-3' Očekávaná velikost amplikonu templátové DNA R. solanacearum: s Rs-1-F/Rs-1-R = 718 bp s Rs-1-F/Rs-3-R = 716 bp
4.2. Reakční směs PCR
a) Protokol PCR specifický pro biovar 1
Reagencie Množství v reakci Konečná koncentrace 0Sterilní UPW 12,925 µl 10x pufr PCR (1) 2,5 µl 1x (1,5 mM MgCl2) BSA (frakce V) (10 %) 0,25 µl 0,1 % Směs dNTP (20mM) 0,125 µl 0,1 mM Primer Rs-1-F (10 µM) 2,0 µl 0,8 µM Primer Rs-1-R (10 µM) 2,0 µl 0,8 µM Taq polymeráza (5U/µl) (1) 0,2 µl 1,0 U Množství vzorku 5,0 µl Celkové množství 25,0 µl
(1) Metoda byla validována použitím Taq polymerázy Perkin Elmer (AmpliTaq)
a Gibco BRL.
b) Protokol PCR specifický pro Biovar 3/4/5
Reagencie Množství v reakci Konečná koncentrace Sterilní UPW 14,925 µl 10x pufr PCR (1) 2,5 µl 1x (1,5 mM MgCl2) BSA (frakce V) (10 %) 0,25 µl 0,1 % Směs dNTP (20mM) 0,125 µl 0,1 mM Primer Rs-1-F (10 µM) 1,0 µl 0,4 µM Primer Rs-3-R (10 µM) 1,0 µl 0,4 µM Taq polymeráza (5U/µl)(1)0,2 µl 1,0 U Množství vzorku 5,0 µl Celkové množství 25,0 µl
----
(1) Metoda byla validována použitím Taq polymerázy Perkin Elmer (AmpliTaq)
a Gibco BRL.
4.3. Reakční podmínky PCR
Provede se následující cyklický proces pro obě reakce specifické pro biovary
1/2- a biovary 3/4/5:
1 cyklus: i) 5 minut při 95 °C (denaturace templátové DNA)
35 cyklů: ii) 30 sekund při 95 °C (denaturace templátové DNA)
iii) 30 sekund při 58 °C (hybridizace primerů s templátovou DNA)
iv) 45 sekund při 72 °C (syntéza kopie)
1 cyklus: v) 5 minut při 72 °C (konečná syntéza)
vi) ponechá se při teplotě 4 °C.
Poznámka:
Tento program byl optimalizován pro použití termocykleru MJ Research
PTC 200. Použití jiných přístrojů může vyžadovat úpravu jednotlivých kroků cyklu
ii), iii) a iv).
4.4. Restrikční enzymová analýza amplikonu.
Produkty PCR vzniklé amplifikací DNA R. solanacearum pomocí primerů Rs-1-F
a Rs-1-R vytvářejí zřetelný polymorfismus délky restrikčních fragmentů s enzymem
Bsm I nebo Isoschizomere (např. Mva 1269 I) po inkubační době 30 minut při teplotě
65 °C. Produkty PCR vzniklé amplifikací z DNA R. solanacearum pomocí primerů Rs-1-F
a Rs-3-R nemají žádná restrikční místa.
5. Příprava nanášecího pufru
5.1. Bromfenolová modř (10 % zásobní roztok)
Bromfenolová modř 5 g Destilovaná (redestilovaná) voda 50 ml
5.2. Nanášecí pufr
Glycerol (86 %) 3,5 ml Bromfenolová modř (5,1) 300 µl Destilovaná (redestilovaná) voda (bidestilát) 6,2 ml
6. Pufr 10x TRIS-acetát-EDTA (TAE), pH 8,0
TRIS 48,4 g Ledová kyselina octová 11,42 ml EDTA (sodná sůl) 3,72 g Destilovaná voda 1,00 l
Před použitím se zředí na 1X.
Také komerčně dostupné (např. Invitrogen nebo rovnocenné).
Dodatek 7
Validovaná činidla pro FISH test
1. Oligosondy
Sonda specifická pro R. solanacearum OLI-1-CY3: 5'-ggc agg tag caa gct acc
ccc-3'
Nespecifická eubakteriální sonda EUB-338-FITC: 5'-gct gcc tcc cgt agg agt-3'
2. Fixační roztok
[UPOZORNĚNÍ: FIXAČNÍ ROZTOK OBSAHUJE PARAFORMALDEHYD, KTERÝ JE TOXICKÝ.
POUŽÍT RUKAVICE A NEVDECHNOUT. DOPORUČUJE SE PRACOVAT V DIGESTOŘI.]
i) Zahřeje se 9 ml molekulárně čisté vody (např. Ultra pure water = (UPW))
na teplotu přibližně 60 °C a přidá se 0,4 g paraformaldehydu. Paraformaldehyd se
rozpustí po přidání 5 kapek 1N NaOH a zamíchání magnetickým míchadlem.
ii) Upraví se pH na 7,0 přidáním 1ml fosfátového pufru 0,1 M (PB; pH 7,0)
a 5 kapek HCl 1N. Zkontroluje se pH indikačním proužkem a v případě potřeby se upraví
pomocí HCl nebo NaOH. [UPOZORNĚNÍ: V ROZTOCÍCH S PARAFORMALEDHYDEM NEPOUŽÍVAT MĚŘIČ
PH!]
iii) Přefiltruje se roztok přes membránový filtr 0,22 µm a skladuje se
chráněný před prachem při teplotě 4 °C do dalšího použití.
3. 3x Hybmix
NaCl 2,7 M Tris-HCl 60 mM (pH 7,4)
EDTA (sterilizovaný přes filtr a autoklávovaný) 15 mM Zředí se až 1x, podle
potřeby.
4. Hybridizační roztok
1x Hybmix Sodium dodecyl sulfát (SDS) 0,01 % Formamid 30 % Sonda EUB 338 5 ng/µl Sonda OLI-1 nebo OLI2 5 ng/µl
Připraví se množství hybridizačního roztoku podle výpočtů v tabulce 1. Pro
každé sklíčko (obsahující dvojmo 2 různé vzorky) je třeba 90 µl hybridizačního roztoku.UPOZORNĚNÍ:
FORMAMID JE VELMI JEDOVATÝ, NUTNO POUŽÍVAT RUKAVICE A UČINIT POTŘEBNÁ BEZPEČNOSTNÍ
OPATŘENÍ!
Tabulka 1. Doporučená množství pro přípravu hybridizační směsi ------------------------------ ---------------------------------------- Počet sklíček 1 4 6 8 10 ------------------------------ ---------------------------------------- Sterilní ultra čistá voda 23,1 92,4 138,6 184,8 231,0 3 x Hybmix 30,0 120,0 180,0 240,0 300,0 1 % SDS 0,9 3,6 5,4 7,2 9,0 Formamid 27,0 108,0 162,0 216,0 270,0 Sonda EUB 338 (100 ng/µl) 4,5 18,0 27,0 36,0 45,0 Sonda OLI-1 nebo OLI2 (100 ng/µl)4,5 18,0 27,0 36,0 45,0 ------------------------------ ---------------------------------------- Celkové množství 90,0 360,0 540,0 720,0 900,0 ------------------------------ ----------------------------------------
Poznámka:
Všechny roztoky obsahující světlocitlivé oligosondy se uchovávají
v temnu při teplotě - 20 °C. Během použití se chrání před přímým slunečním zářením
nebo elektrickým světlem.
5. 0,1M fosfátový pufr, pH 7,0
Na2HPO 48,52 g KH2PO45,44 g Destilovaná voda 1,00 l
Rozpustí se složky, zkontroluje pH a sterilizuje autoklávováním při teplotě
121 °C po dobu 15 minut.
Dodatek 8
Pěstování lilku a rajčete
Vysejí se semena rajčete (Lycopersicon esculentum) nebo lilku (Solanum melongena)
do pasterizovaného výsevního substrátu. Sazeničky s plně rozvinutými děložními lístky
(10 až 14 dní) se přesadí do pasterizovaného pěstebního substrátu. Rostliny lilku
a rajčete by se měly pěstovat ve skleníku za následujících podmínek:
Délka dne: 14 hodin nebo přirozená délka dne, pokud je delší;
Teplota: den: 21 až 24 °C,
noc: 14 až 18 °C.
Vhodná odrůda lilku vejcoplodého: "Black Beauty"
Vhodná odrůda rajčete: "Moneymaker"
Dodavatelé: viz internetová stránka http://forum.europa.eu.int/Public/irc/sanco/Home/main.
LITERATURA
1. Amann, R.I., L. Krumholz and D.A. Stahl. 1990. Fluorescent-oligonucleotide
probing of whole cells for determinative, phylogenetic and environmental studies
in microbiology. J. Bacteriol. 172: 762-770.
2. Anon. 1998. Council Directive 98/57/EC of 20 July 1998 on the control
of Ralstonia solanacearum (Smith) Yabuuchi et al. Official Journal of the European
Communities L235, 1-39.
3. Boudazin, G., A.C. Le Roux, K. Josi, P. Labarre and B. Jouan. 1999.
Design of division specific primers of Ralstonia solanacearum and application to
the identification of European isolates. European Journal of Plant Pathology 105;373-380.
4. Caruso, P., Gorris, M.T., Cambra, M., Palomo, J.L., Collar, J and Lopez,
M.M. 2002. Enrichment Double-Antibody Sandwich Indirect Enzyme-Linked Immunosorbent
Assay That Uses a Specific Monoclonal Antibody for sensitive Detection of Ralstonia
solanacearum in Asymptomatic Potato Tubers. Applied and Environmental Microbiology,
68, 3634-3638.
5. Cook, D., Barlow, E. and Sequeira, L. 1989. Genetic diversity of Pseudomonas
solanacearum: detection of restriction fragment length polymorphisms with DNA probes
that specify virulence and the hypersensitive response. Molecular Plant-Microbe Interactions
1:113-121.
6. Elphinstone, J.G., Hennessy, J., Wilson, J.K. and Stead, D.E. 1996.
Sensitivity of detection of Ralstonia solanacearum in potato tuber extracts. EPPO
Bulletin 26; 663- 678.
7. Englebrecht, M.C. (1994) Modification of a semi-selective medium for
the isolation and quantification of Pseudomonas solanacearum. In: A.C. Hayward (ed.)
Bacterial Wilt Newsletter 10, 3-5. Australian Centre for International Agricultural
Research, Canberra, Australia.
8. Hayward, A.C. 1964. Characteristics of Pseudomonas solanacearum. Journal
of Applied Bacteriology 27; 265-277.
9. Hayward, A.C., El-Nashaar, H.M., Nydegger, U. and De Lindo, L. 1990.
Variation in nitrate metabolism in biovars of Pseudomonas solanacearum. Journal of
Applied Bacteriology 69; 269-280.
10. Ito, S., Y. Ushijima, T. Fujii, S. Tanaka, M. Kameya- Iwaki, S. Yoshiwara
and F. Kishi. 1998. Detection of viable cells of Ralstonia solanacearum in soil using
a semi-selective medium and a PCR technique. J. Phytopathology 146; 379-384.
11. Janse, J.D. (1988) A detection method for Pseudomonas solanacearum
in symptomless potato tubers and some data on its sensitivity and specificity. Bulletin
OEPP/EPPO Bulletin 18, 343-351.
12. Janse, J.D. 1991. Infra- and intra-specific classification of Pseudomonas
solanacaerum strains using whole cell fatty-acid analysis. Systematic and Applied
Microbiology 14; 335-345.
13. Kelman, A. 1954. The relationship of pathogenicity of Pseudomonas solanacearum
to colony appearance on a tetrazolium medium. Phytopathology 44; 693-695.
14. Klement Z.; Rudolph, K and D.C. Sands, 1990. Methods in Phytobacteriology.
Akadémiai Kiadó, Budapest, 568 pp.
15. Lelliott, R.A. and Stead, D.E. 1987. Methods for the diagnosis of bacterial
diseases of plants. Blackwell scientific Publications Ltd., Oxford. 216 pp.
16. Lopez, M.M., Gorris, M.T., Llop, P., Cubero, J., Vicedo, B., Cambra,
M., 1997. Selective enrichment improves selective isolation, serological and molecular
detection of plant pathogenic bacteria. In: H.W. Dehne et al., (eds). Klewer Academic
Publishers. pp. 117-121.
17. Louws, F.J., Fulbright, D.W., Stephens, C.T. and De Bruijn, F.J., 1994.
Specific genomic fingerprints of phytopathogenic Xanthomonas and Pseudomonas pathovars
and strains generated with repetitive sequences and PCR. Applied and Environmental
Microbiology, 60, 2286-2295.
18. Louws, F.J., Fulbright, D.W., Stephens, C.T. and De Bruijn, F.J. 1995.
Differentiation of genomic structure by rep- PCR fingerprinting to rapidly classify
Xanthomonas campestris pv. vesicatoria. Phytopathology 85; 528-536.
20. Pastrik, K.H. and Maiss, E. 2000. Detection of R. solanacearum in potato
tubers by polymerase chain reaction. J. Phytopathology 148; 619-626.
19. Opina, N., F. Tavner, G. Holloway, J.-F Wang, T.-H Li, R. Maghirang,
M. Fegan, A.C. Hayward, V. Krishnapillai, W.F. Hong, B.W. Holloway, J.N. Timmis.
1997. A novel method for development of species and strain-specific DNA probes and
PCR primers for identifying Burkholderia solanacearum (formerly Pseudomonas solanacearum).
As Pac. J. Mol. Biol. Biotechnol. 5; 19-33.
21. Pastrik, K.H., Elphinstone, J.G. and Pukall, R. 2002. Sequence analysis
and detection of Ralstonia solanacearum by multiplex PCR amplification of 16S-23S
ribosomal intergenic spacer region with internal positive control. European Journal
of Plant Pathology 108, 831-842.
22. Robinson-Smith, A., Jones, P., Elphinstone, J.G. and Forde, S.M.D.
(1995) Production of antibodies to Pseudomonas solanacearum, the causative agent
of bacterial wilt. Food and Agricultural Immunology 7, 67-79.
23. Schaad, W. 2001. Laboratory guide for identification of plant pathogenic
bacteria. Schaad [Hrsg.]. - 3. ed.; St. Paul, Minnesota: 373 pp.
24. Seal, S.E., L.A. Jackson, J.P.W. Young, and M.J. Daniels. 1993. Detection
of Pseudomonas solanacearum, Pseudomonas syzygii, Pseudomonas pickettii and Blood
Disease Bacterium by partial 16S rRNA sequencing: construction of oligonucleotide
primers for sensitive detection by polymerase chain reaction. J. Gen. Microbiol.
139: 1587-1594.
25. Smith, J.J., Offord, L.C., Holderness, M. and Saddler, G.S. 1995. Genetic
diversity of Burkholderia solanacearum (synonym Pseudomonas solanacearum) race 3
in Kenya. Applied and Environmental Microbiology 61; 4262-4268.
26. Stead, D.E. 1992. Grouping of plant pathogenic and some other Pseudomonas
spp. using cellular fatty-acid profiles. International Journal of Systematic Bacteriology
42; 281-295.
27. Taghavi, M., Hayward, A.C., Sly, L.I., Fegan, M. 1996. Analysiss of
the phylogenetic relationships of strains of Burkholderia solanacearum, Pseudomonas
syzygii, and the blood disease bacterium of banana based on 16S rRNA gene sequences.
International Journal of Systematic Bacteriology 46; 10-15.
28. Van Der Wolf, J.M., Bonants, P.J.M., Smith, J.J., Hagenaar, M., Nijhuis,
E., Van Beckhoven, J.R.C., Saddler, G.S., Trigalet, A., Feuillade, R. 1998. Genetic
diversity of Ralstonia solanacearum Race 3 in Western Europe as determined by AFLP,
RC-PFGE and rep-PCR. In: Prior, P., Allen, C. and Elphinstone, J. (eds.) Bacterial
wilt disease: Molecular and Ecological Aspects. Springer (Berlin) pp. 44-49.
29. Weller, S.A., Elphinstone, J.G., Smith, N., Stead, D.E. and Boonham,
N. 1999. Detection of Ralstonia solanacearum strains using an automated and quantitative
flourogenic 5' nuclease TaqMan assay. Applied and Environmental Microbiology 66;
2853-2858.
30. Wullings, B.A., A.R. van Beuningen, J.D. Janse and A.D.L. Akkermans.
1998. Detection of R. solanacearum, which causes brown rot of potato, by fluorescent
in situ hybridization with 23s rRNA-targeted probes. Appl. Environ. Microbiol. 64:
4546-4554.